
Cet article fait suite à notre article intitulé Thermodynamique et sens de la vie. Il est recommandé de lire la suite d’articles dans l’ordre.
Une niche occupée peut être considérée comme une interdiction pour une autre espèce de l’occuper. Il est parfois plus opportun de remplacer ce terme (interdiction) par celui de “contraintes”, qui peut sembler moins catégorique. Bien entendu, outre les contraintes liées à la disponibilité ou à l’absence de ressources, il existe d’autres types de contraintes, telles que les contraintes thermodynamiques, climatiques ou édaphiques. Leur impact sur l’évolution est bien connu et nous allons également les aborder. En substance, l’interdiction peut être associée au fait que toutes les trajectoires évolutives ne garantissent pas la stabilité des systèmes vivants. Par exemple, les contraintes au niveau de l’écosystème sont avant tout des rétroactions négatives, qui ne permettent pas aux espèces d’évoluer de manière à perturber le cycle local des nutriments. Il est donc possible de considérer les permissions comme des rétroactions positives et les interdictions comme des rétroactions négatives.
Edmundas Lekevičius, Ecological Darwinism or Preliminary Answers to Some Crucial though Seldom Asked Questions
On peut donc identifier un sens à la vie grâce à deux grands principes connus à ce jour qui sont la thermodynamique et l’évolution. Il n’est pas impossible que l’on en découvre plus dans le futur et des travaux établissant un lien avec la physique quantique commencent déjà à porter leurs fruits, mais laissons cela de côté pour l’instant. Si la thermodynamique est un principe global qui impacte tout l’univers, l’évolution darwinienne va, elle, créer les particularités du vivant.
On peut alors appréhender la vie sous un autre angle et la définir selon une approche darwinienne comme un système moléculaire autoreproducteur disposant de qualités que sont la capacité à métaboliser, se reproduire et évoluer, ou en d’autres termes, un système chimique auto-entretenu capable d’évolution.
De ce point de vue, le but d’un organisme sera de passer ses gènes à la génération suivante. Si la thermodynamique nous explique que la fin de l’univers est de finir dans un tout, un et indifférencié à cause de l’entropie inéluctable, l’évolution semble pourtant au contraire générer des systèmes extropiques. Mais est-ce que l’évolution est conservatrice ou productive ? C’est-à-dire, est-ce qu’elle ne permet que l’adaptation à un environnement donné ou pousse-t-elle naturellement à un dépassement ?
L’évolution comme un algorithme
On peut voir l’évolution comme un algorithme. Elle peut être considérée comme un mécanisme d’extraction de régularités statistiques, une sorte d’algorithme de renforcement opérant sur l’échelle de temps des générations, où le signal de renforcement consiste en le nombre de descendants qu’un individu génère qui est directement lié à sa capacité à survivre et à se reproduire.
Nos sens ne sont alors que des signaux que l’évolution a sélectionnés, car ils nous offrent une façon plus optimisée d’appréhender le monde. La fonction du plaisir va nous indiquer ce qui est bon pour nous, et donc qui sert la vie et favorise la transmission de nos gènes alors que les sensations d’inconfort et de dégoût vont nous indiquer ce que nous devons fuir. On va chercher à se reproduire avant tout parce qu’on trouve une sensation de plaisir dans l’acte sexuel.
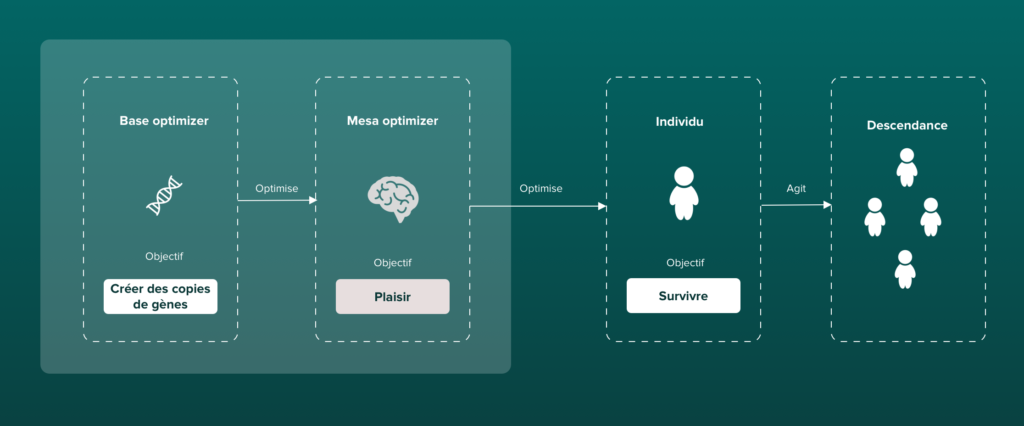
Il est possible de dresser un parallèle entre le fonctionnement de l’évolution et celui du Deep Learning, une branche de l’intelligence artificielle. Imaginez un système qui apprend et s’adapte constamment à son environnement, tout comme une créature vivante. C’est essentiellement ce que fait le “Deep Learning”. Il utilise des réseaux de neurones artificiels pour apprendre des schémas complexes à partir de données et faire des prédictions. Mais comment cela fonctionne-t-il exactement et en quoi cela ressemble-t-il à l’évolution ?
En fait, le deep learning est un peu comme un écosystème numérique. Chaque “neurone” dans le réseau est comme une petite créature qui essaie de s’adapter à son environnement. Ce “neurone” ajuste ses “poids” ou paramètres pour mieux correspondre aux données qu’il reçoit, un peu comme une créature qui évolue pour mieux survivre dans son environnement.
Cette idée nous vient du concept évoqué précédemment d’adaptation dissipative proposé par le physicien Jeremy England. Les structures qui sont capables de bien utiliser l’énergie de leur environnement ont tendance à persister et à se reproduire. Dans le contexte du deep learning, les “neurones” qui peuvent bien correspondre aux données – ou “observations” – qu’ils reçoivent sont plus susceptibles de rester dans le réseau.
Maintenant, vous pourriez penser que tout cela ressemble beaucoup à de l’optimisation – après tout, les “neurones” essaient de “minimiser” leur erreur. Mais il y a une différence importante. Dans l’optimisation classique, nous cherchons le meilleur résultat possible, tandis que dans le deep learning, nous cherchons à “généraliser” à partir des données, ce qui signifie être capable de faire de bonnes prédictions même sur des données nouvelles et inconnues.
Pour continuer avec notre analogie, c’est un peu comme si une créature s’adaptait non seulement à son environnement actuel, mais aussi à de nouveaux environnements qu’elle n’a jamais rencontrés. C’est un défi beaucoup plus grand, mais c’est exactement ce que le deep learning réussit à faire.
L’évolution, tout comme le Deep Learning, est un processus productif et non conservateur. Cela signifie qu’elle ne se contente pas de maintenir un état ou une configuration existante, mais qu’elle génère continuellement de nouvelles configurations en réponse à son environnement.
Dans le cas du deep learning, ces nouvelles configurations sont représentées par les poids ajustés des neurones du réseau, qui changent en réponse aux données d’entrée. De la même manière, l’évolution produit de nouvelles variations génétiques et des traits adaptatifs en réponse aux pressions de l’environnement.
Ces processus sont donc fondamentalement dynamiques et adaptatifs. Ils sont régis par des principes d’auto-organisation, qui favorisent les configurations qui sont en harmonie avec leur environnement et qui peuvent efficacement dissiper l’énergie ou minimiser l’entropie.
But de l’évolution et cybernétique
Est-ce que le sens de la vie est la production d’entropie ? En-tout-cas, si ce n’est pas son but final, c’est une condition qui guide cette dernière et la meilleure façon de produire de l’entropie est de mémoriser de l’information afin de la convertir en énergie libre à dissiper dans un travail mécanique. La dissipation d’énergie étant directement liée à la diminution d’entropie du système, c’est-à-dire à l’information mémorisée, ledit système s’auto-organise en mémorisant de l’information sur son environnement. Il mémorise l’information qui lui permet de maximiser la production d’énergie libre. Il est donc clair que, du point de vue thermodynamique, le processus de sélection naturelle s’applique non pas aux structures elles-mêmes, mais à l’information qu’elles mémorisent.
Dans le cas des plantes ou des animaux, elle s’applique donc aux gènes, mais dans le cas des sociétés humaines, elle s’appliquerait ainsi sur la capacité à mémoriser et utiliser de l’information afin de produire de l’énergie libre convertie en travail mécanique. Plus un organisme, une espèce et une civilisation mémorisent d’information, plus elles dissipent d’énergie.
« La sélection naturelle favorise l’organisme qui dissipe le plus vite l’énergie. »
– Alfred Lotka, Natural selection as a physical principle
« La lutte pour la vie est une lutte pour dissiper l’énergie. »
– Ludwig Boltzmann, Popular writings
Ainsi, le sens de l’évolution allant de LUCA (Last Universal Common Ancestor) à nos jours, qui a vu naître homo-sapiens et la civilisation, est de sélectionner les structures qui vont stocker de mieux en mieux l’information, produire de plus en plus de connaissances et consommer de plus en plus d’énergie. L’évolution peut alors être appréhendée comme un système cybernétique constitué de circuits de rétroaction adaptatifs. On ne peut pleinement la comprendre sans la penser sous l’angle de la théorie de l’information de Claude Shannon et de la cybernétique de Wiener et sans comprendre qu’elle maximise une valeur impliquant la dissipation d’énergie. Mais comment fonctionne-t-elle concrètement dans nos interactions entre individus ?
Les deux mécanismes de la sélection naturelle
Dans l’inconscient collectif, le terme de sélection naturelle renvoie à la lutte pour la survie. En réalité, la sélection naturelle accompagne un deuxième mécanisme qui est la sélection sexuelle.
Pour le dire simplement, on va chercher à éviter les dangers et la souffrance afin d’augmenter nos chances de survie, mais aussi optimiser notre plaisir, notamment dans l’activité sexuelle. On sera alors en compétition entre espèces pour la survie ainsi qu’au sein même de notre espèce pour se reproduire. Ces deux phénomènes vont alors exercer une pression de sélection.
On va observer un compromis entre la lutte pour la survie et la lutte pour la reproduction. Il arrive parfois que des traits se révèlent avantageux pour l’un, mais pas pour l’autre. Par exemple, le plumage de parade nuptiale du paon est fort apprécié par le sexe opposé, mais il lui rend la tâche plus délicate pour échapper aux prédateurs.
La sélection sexuelle repose alors elle-même sur deux mécanismes ; la sélection intersexuelle qui va impacter le choix du partenaire – les femelles choisissant les mâles dans la majorité des espèces – et la sélection intrasexuelle qui génère la compétition pour l’accès aux membres du sexe opposé, qu’on observe aussi bien chez les mâles que chez les femelles, et qui va s’exprimer de façons différentes.
La compétition intrasexuelle se réfère à la compétition entre individus du même sexe, souvent pour l’accès aux partenaires. Chez les hommes, cette compétition peut souvent prendre des formes plus directes et physiques, en raison de l’héritage de notre passé évolutif où la force physique était un déterminant majeur de l’accès aux ressources et aux partenaires. Les femmes, en revanche, peuvent être plus susceptibles d’utiliser des stratégies de compétition plus indirectes et sociales, telles que l’exclusion sociale ou la manipulation de la réputation. Cette double compétition intergroupe et intragroupe va se retrouver au-delà des sexes et va naturellement conduire à différents niveaux de sélections.
Les différents niveaux de sélection
« L’égoïsme supplante l’altruisme au sein d’un groupe. Les groupes altruistes supplantent les groupes égoïstes. Tout le reste n’est que commentaire. »
– E.O Wilson
L’évolution influence aussi bien nos comportements individuels au sein de notre groupe que celui de notre groupe parmi les autres groupes. Elle repose sur plusieurs niveaux de sélection s’appliquant aux gènes, à l’individu et au groupe. Ces différents niveaux s’imbriquent comme des poupées russes et vont s’influencer l’un l’autre au travers de différents procédés.
Une explication exhaustive demanderait un livre entier mais je vais tenter de résumer le procédé brièvement en me focalisant sur les points clés qui influencent nos interactions avec autrui. Comme l’a popularisé Richard Dawkins dans son livre Le gène égoïste, la première des sélections s’effectue au niveau génétique. Contrairement à l’individu qui est mortel, le gène est lui “immortel” et se sert de l’individu comme “véhicule”. Le gène se réplique afin d’assurer sa survie et se transmettre de génération en génération via la reproduction de l’individu.
Pour reprendre des termes cybernétiques, chez les humains pratiquant la reproduction sexuée, on observe une première boucle négative qui va assurer une certaine stabilité en favorisant la reproduction entre petits cousins – deux individus ayant en commun un couple d’arrière-arrière-grands-parents. Les unions consanguines présentant trop peu de brassage génétique sont pénalisées en rendant souvent la progéniture stérile. Mais, phénomène moins connu et pourtant logique, l’union entre des organismes très différents, que l’on nomme la dépression hybride, ne constitue pas un avantage sélectif. Les unions entre deux organismes trop éloignés génétiquement sont tout simplement rendus impossibles par le phénomène de spéciation ou donnent, dans le meilleur des cas, des descendances moins nombreuses, et dans le pire des cas, des progénitures là aussi stériles, comme le mulet chez les animaux.
La transmission d’un grand nombre de copies de ses gènes s’effectue donc le plus directement en maximisant la reproduction. Le comportement n’est qu’un épiphénomène, un moyen de transmettre des copies de gènes à la génération suivante et la sélection individuelle réussit mieux que la sélection de groupe à expliquer les comportements de base. Imaginez qu’une hyène s’abatte sur un groupe de zèbres. Que ferait le plus proche si la sélection s’effectue uniquement sur le groupe ? Il resterait là, se sacrifiant pour le groupe. En revanche, si la sélection individuelle prime alors le zèbre courrait comme un dératé… et les zèbres courent comme des dératés. Prenons maintenant les hyènes qui viennent de tuer le zèbre. Dans un cadre de sélection de groupe, tout le monde mangerait calmement à tour de rôle. Si la sélection était individuelle alors on assisterait à une frénésie générale… Bingo ! C’est ce qui se produit.
Mais les individus ont des gènes en commun avec d’autres individus. 50% de gènes en commun avec leurs frères et sœurs (100% en cas de jumeaux homozygotes), 25% avec leurs cousins etc. La sélection s’effectuant en premier lieu sur une base génétique, les individus vont naturellement adopter un comportement différent selon la proximité génétique des gens avec qui ils interagissent, le but étant de favoriser la reproduction de leurs gènes portés par les individus qui leur sont proches. On appelle cela la sélection de parentèle et elle fut découverte par Hamilton qui proposa pour la décrire un modèle mathématique sophistiqué.
En termes mathématiques, cela donne l’équation [rB > C], où [B] indique les bénéfices (benefits) du comportement altruiste, [C] (cost) leur coût et [r] la corrélation génétique (relatedness) entre l’acteur et le bénéficiaire de son comportement.
On observe un comportement altruiste entre individus ayant un fort taux d’apparentement, c’est-à-dire quand les bénéfices du receveur sont supérieurs au coût pour le donneur. Nous préférons coopérer avec les individus partageant le plus de gènes en commun car cela permet de favoriser la propagation de nos gènes. L’individu cherche à transmettre ses propres allèles, que ce soit par l’intermédiaire de sa propre reproduction ou par celle des individus qui lui sont apparentés.
Cela va donc expliquer les comportements altruistes qui eux ont un impact sur le groupe. Un comportement altruiste traduit la volonté d’un individu au sein d’un groupe à réduire sa propre aptitude (ou capacité d’un individu à se reproduire dans un environnement et un contexte donné) pour le bien de celle d’un ou plusieurs autres individus du groupe. Ce trait sera sélectionné et transmis aux générations suivantes car il permet au groupe d’avoir une meilleure aptitude. On peut parler ici de conflit groupe-individu car un trait désavantageux au niveau de l’individu (l’altruisme, qui réduit l’aptitude de l’individu) est pourtant sélectionné puisqu’il rapporte une meilleure aptitude au niveau du groupe. Il n’y a cependant pas toujours conflit et un trait favorable au groupe peut aussi être favorable à l’individu.
Cependant, l’altruisme ne peut pas toujours être justifié par la seule sélection de parentèle. En plus de ces niveaux de sélection, on observe des procédés existants entre individus ne possédant pas ou très peu de gènes en commun, et c’est ici que va intervenir le plus la raison.
Pour expliquer pourquoi des individus s’entraident même sans lien de parenté, Robert Trivers a introduit le concept d’altruisme réciproque. Prenons l’exemple d’une personne qui risque sa vie pour sauver un inconnu de la noyade. Si cet acte se produit isolément, il semble désavantageux. Cependant, si ces actes de sauvetage sont réciproques, les individus impliqués bénéficient d’une augmentation de leurs chances de survie. Dans une population où la noyade est un risque courant, ceux qui pratiquent l’altruisme réciproque ont tendance à avoir de meilleures chances de survie et de reproduction que ceux qui ne le font pas.
Robert Trivers met en évidence que l’altruisme réciproque est fréquent chez les humains, englobant des actions telles que le secours en cas de danger, le partage de ressources, et l’aide aux personnes vulnérables. Au sein de l’espèce humaine, l’altruisme réciproque est plus complexe que chez d’autres espèces en raison de la conscience et du langage. Les individus peuvent évaluer la possibilité d’une réciprocité ou choisir de tromper. De plus, des émotions telles que l’indignation, la honte, et la culpabilité jouent un rôle crucial dans la formation de relations durables en dehors de la famille et semblent avoir évolué pour favoriser l’altruisme et la réciprocité en décourageant la tromperie et l’égoïsme.
Richard D. Alexander a proposé dans les années 1980 une hypothèse complémentaire à l’altruisme réciproque, nommée “réciprocité indirecte”. Elle explique la coopération dans les grandes sociétés où la sélection de parentèle et l’altruisme réciproque ne sont pas toujours applicables. Dans la réciprocité indirecte, un individu A aide un individu B (sans lien de parenté), et en retour, reçoit de l’aide d’un individu C (également sans lien de parenté). Cela est basé sur le fait qu’être altruiste peut améliorer la réputation d’un individu, ce qui favorise des relations de confiance et des bénéfices à long terme. La stratégie optimale pour maximiser les bénéfices de tous est de coopérer constamment, sauf si l’autre partie ne coopère pas, ce qui est connu sous le nom de “tit for tat”, tirée de la Théorie des jeux formulée par John Von Neumann qui est tellement un génie qu’il est difficile de le présenter sous un seul angle.
Enfin, il existe aussi un phénomène appelé la pseudo-parenté (pseudokinship) qui consiste en la manipulation des sentiments d’un être vivant pour qu’il se considère plus proche d’un autre qu’il ne l’est réellement. C’est là par exemple qu’interviennent les idéologies, religions ou aires culturelles que créent les empires. L’appartenance des individus à ces entités ne repose pas nécessairement sur leur lien de parenté et pourtant, ils se sentent frères. Le pendant de la pseudo-parenté se nomme pseudo-spéciation, il s’agit à l’inverse de la manipulation des sentiments d’un être vivant pour qu’il se considère plus éloigné d’un autre qu’il ne l’est réellement. Cela s’illustre notamment par notre tendance à déshumaniser les membres d’un autre groupe en les faisant passer pour des animaux, rats ou agents infectieux. Cela ne remplace pas la sélection de parentèle “classique” mais cela ajoute un degré de complexité à son mécanisme.
L’évolution comme principe global
Dissiper l’énergie et mémoriser l’information sont exactement la même chose. Ce qui compte, c’est donc l’information et non le gène. Le gène est simplement un moyen de mémoriser de l’information sur son environnement afin de dissiper l’énergie le plus efficacement possible.
Or, le gène n’est pas le moyen le plus efficace de mémoriser l’information. Depuis la révolution cognitive ayant eu lieu autour de -70,000, le cerveau de l’homme s’est hypertrophié lui conférant des capacités supérieures dans ce domaine. Ses fonctions cognitives se sont grandement améliorées et avec elles, le langage et plus globalement la communication. Du gène, nous sommes passés à ce que Dawkins appelle le mème, qu’il conçoit comme la plus petite unité d’information que l’on échange via la culture.
Dès lors, la culture va devenir un élément extrêmement important des sociétés humaines. On soupçonne que les mèmes sont soumis aux mêmes principes de l’évolution et je reviendrai plus en détails sur le sujet des gènes et des mèmes plus tard mais la base scientifique du mème est discutable donc je m’en tiendrais à cela pour l’instant. On peut cependant penser l’évolution de façon globale comme un va et vient entre une Macroévolution et une Microévolution. Il y aurait une alternance de macro et de micro évolution de plus en plus fréquente depuis le Big Bang.
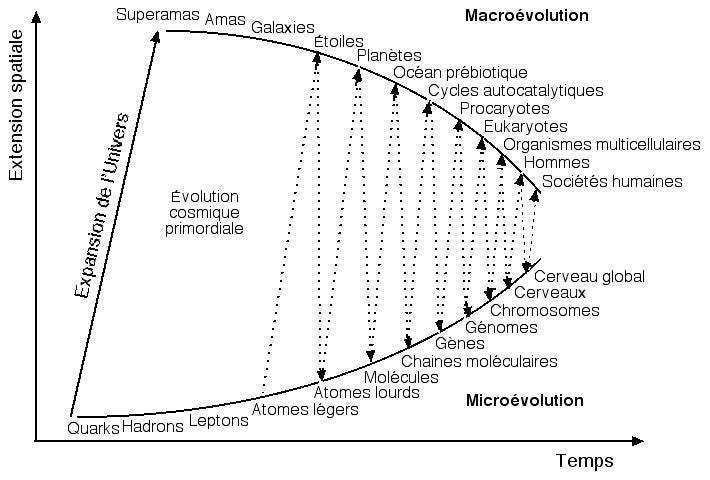
L’univers qui s’auto organise, d’après Eric Jantsh et retravaillé par François Roddier. On ne sait pas ce qu’il se passe aux tout premiers stades après le Big Bang. Mais depuis les atomes légers, on ne cesse d’observer une alternance entre une micro et une macro évolution.
La notion d’évolution doit alors être comprise de façon globale et dépasser la simple évolution darwinienne qui se borne aux phénomènes observables sur Terre. Toute évolution d’un système est soumise au contrôle d’un méta-système. L’évolution darwinienne doit être comprise comme un ensemble de processus au service de la biosphère qui forme le méta-système.
Valentin Turchin, cybernéticien et pionnier de l’informatique a travaillé sur le concept de “meta-system transition” (MST) dans le cadre de sa théorie cybernétique de l’évolution, qui envisage l’évolution comme une série de transitions vers des niveaux de complexité et d’intégration de plus en plus élevés. Chaque MST représente une étape où un ensemble de systèmes se regroupe et forme un méta-système plus complexe. Chaque nouveau “meta-système” représente un niveau d’organisation plus élevé effectuant un contrôle sur les sous-systèmes. Dans cette perspective, les entités simples s’associent pour former des structures plus complexes, qui à leur tour peuvent s’associer pour former des niveaux encore plus élevés d’organisation.
Il est alors fort logique de comprendre pourquoi un méta-système va favoriser l’altruisme de ses différentes parties, mais dans le même temps être lui-même égoïste. Chaque système étant inclus dans un méta-système, il devra constamment trouver un compromis entre ses désirs égoïstes et les besoins altruistes dictés par le méta-système. L’altruisme concernera évidemment les membres de son groupe, mais même toute entité appartenant au système comme des animaux appartenant à d’autres espèces. On peut inclure dans cela tous les processus de domestications animalières comme celles du chien et du cheval.
En appliquant le concept de MST à la biosphère, qui englobe tous les écosystèmes de la Terre, on peut comprendre comment l’évolution darwinienne est soumise aux conditions de ce vaste méta-système. Les individus et les populations ne sont pas isolés ; ils interagissent continuellement avec d’autres organismes et avec leur environnement. Ces interactions peuvent aboutir à la formation de systèmes plus complexes, tels que des communautés d’espèces ou des écosystèmes.
Dans ce contexte, la sélection naturelle peut agir non seulement au niveau de l’individu, mais aussi à des niveaux supérieurs d’organisation. Par exemple, certains traits ou comportements peuvent être favorisés parce qu’ils contribuent à la stabilité ou au succès d’une communauté ou d’un écosystème, et pas seulement à la réussite reproductive de l’individu.
Le concept de méta-systèmes de transition de Turchin élargit alors notre compréhension de l’évolution darwinienne en montrant comment elle est intégrée dans des hiérarchies de systèmes complexes. Cela met en lumière l’importance des interactions et des dynamiques à différents niveaux d’organisation, et comment elles peuvent influencer le cours de l’évolution au sein de la biosphère.
« Que se passerait-il dans la situation inverse, c’est-à-dire si ces relations étaient uniquement de type compétitif ? Je pense que le résultat final le plus probable serait qu’une seule espèce existerait dans un endroit donné à un moment donné ; c’est-à-dire celle qui aurait remplacé toutes les autres espèces, celles qui ne sont pas si bien adaptées à la lutte pour l’existence. Et au sein de cette espèce, il n’existerait qu’un seul génotype (“sauvage”) ayant remplacé tous les génotypes de moindre valeur adaptative. Naturellement, il n’y aurait pas de communautés écologiques ou d’écosystèmes dans ces circonstances. Cependant, comme cela a déjà été mentionné, ce type de vie n’aurait aucune chance de survie puisqu’aucune des espèces ne peut maintenir un cycle de nutriments à elle seule. En résumé, on peut affirmer que la vie a choisi un compromis entre deux stratégies incompatibles : être le plus efficace possible et ne pas compromettre l’adaptabilité. Ce compromis a dû conditionner l’existence à long terme de la vie. Cependant, le problème de ce type de réponse réside dans ses implications téléologiques. Nous pourrions parvenir à une bien meilleure réponse si nous découvrions les processus évolutifs par lesquels cette forme de vie a pu émerger. »
– Edmundas Lekevičius, Ecological Darwinism or Preliminary Answers to Some Crucial though Seldom asked Questions
Dionysien et Apollinien
Il est tout à fait possible de concevoir la thermodynamique et l’évolution comme des forces respectivement dionysienne et apollinienne. La tragédie nous révèle explicitement la façon de penser des Grecs et tout spécialement des présocratiques. La tragédie grecque est composée d’un chœur de danseurs dionysiens qui vont entourer la scène et s’agiter tandis que les acteurs prennent place et jouent leur rôle au sein de cette dernière, finissant inéluctablement dans la mort. Si le chœur représente le Dionysien, les acteurs incarnent eux la représentation apollinienne, l’individuation de l’Un-primordial en différentes entités qui apparaissent à partir du Dionysien avant d’y retourner.
De la même façon, on peut tout à fait voir la thermodynamique comme le principe qui va faire émerger la vie, avant que l’évolution ne la sculpte en différentes entités, la fin étant la mort thermique de l’univers et de ces individuations. En rapportant cela à l’information, on peut voir Dionysos comme le chaos ou l’information non structurée qui cherche encore un ordre et Apollon comme l’information ordonnée qui va porter un individu à l’existence.
Cet article vous a plus ? Soutenez nous sur Tipeee




