
Une étude vient de paraître ayant potentiellement de vastes implications pour comprendre ce qui a permis à Sapiens de déplacer les Néandertaliens et d’autres espèces d’Homo dans le passé géologiquement récent, ainsi que de remodeler littéralement le monde au cours de l’Anthropocène. Elle met en avant des différences génétiques potentiellement liées à l’essor de la créativité chez Homo-Sapiens qui n’existe que partiellement chez Néandertal et de façon encore plus parcellaire chez les chimpanzés. Cette étude nous semblant extrêmement intéressante, le contenu que vous trouverez ci-dessous en est une traduction fidèle, donc longue.
La base génétique de l’émergence de la créativité chez l’homme moderne reste un mystère malgré le séquençage des génomes des chimpanzés et des Néandertaliens, nos plus proches parents hominidés. Des méthodes fondées sur des données nous ont permis de découvrir des réseaux de gènes qui distinguent les trois principaux systèmes de la personnalité et de l’adaptabilité de l’homme moderne : la réactivité émotionnelle, la maîtrise de soi et la conscience de soi. Nous avons maintenant identifié lesquels de ces gènes sont présents chez les chimpanzés et les Néandertaliens. Nous avons reproduit nos résultats dans des analyses distinctes de trois génomes de Néandertaliens. Nous avons constaté que les Néandertaliens possédaient presque les mêmes gènes de réactivité émotionnelle que les chimpanzés, et qu’ils se situaient à un niveau intermédiaire entre les humains modernes et les chimpanzés en ce qui concerne le nombre de gènes de maîtrise de soi et de conscience de soi. 95 % des 267 gènes que nous avons trouvés uniquement chez l’homme moderne ne codaient pas de protéines, y compris de nombreux ARN non codants dans le réseau de la conscience de soi. Ces gènes peuvent être le résultat d’une sélection positive pour les caractéristiques du bien-être humain et de la modernité comportementale, notamment la créativité, le comportement prosocial et la longévité. Les gènes qui se regroupent en association avec ceux que l’on ne trouve que chez l’homme moderne sont surexprimés dans les régions du cerveau impliquées dans la conscience de soi et la créativité chez l’homme, y compris les régions du néocortex à myélinisation tardive et phylogénétiquement récentes pour la mémoire autobiographique dans les régions frontales, pariétales et temporales, ainsi que les composants connexes des boucles cortico-thalamo-ponto-cérébelleuses-corticales et cortico-striato-corticales. Nous concluons que l’homme moderne possède plus de 200 gènes uniques non codants pour les protéines qui régulent la co-expression de beaucoup plus de gènes codants pour les protéines dans des réseaux coordonnés qui sous-tendent ses capacités de conscience de soi, de créativité, de comportement prosocial et de longévité en bonne santé, qui ne se retrouvent pas chez les chimpanzés ou les Néandertaliens.
Traité
Néoréactionnaire
Le premier livre de NIMH
Traité
Néoréactionnaire
Le premier livre de NIMH
Introduction
L’une des questions les plus fondamentales sur la nature humaine est de savoir ce qui a déclenché l’émergence explosive de la créativité chez les humains modernes avant leur dispersion à grande échelle depuis l’Afrique et l’extinction consécutive des Néandertaliens [1,2,3,4]. D’importantes controverses persistent sur les fondements de la créativité humaine en art et en science, ainsi que sur les différences de cognition, de langage et de personnalité qui distinguent les humains modernes des hominidés disparus [5,6,7,8]. Ces controverses sont dues au fait que les archives archéologiques et fossiles sont incomplètes et sujettes à une interprétation ambiguë [9, 10].
Qu’est-ce qui distingue les humains modernes sur le plan comportemental des autres hominidés ?
Les anthropologues distinguent l’Homo sapiens (Sapiens) comportementalement moderne des autres hominidés en raison de l’adaptabilité cognitive, sociale et physique accrue de Sapiens. Les Sapiens modernes du point de vue comportemental font preuve d’une créativité remarquable par rapport aux autres hominidés : ils montrent des signes d’innovation, de flexibilité, de profondeur de planification et de capacités cognitives liées au symbolisme et à la conscience de soi qui permettent la génération spontanée d’art narratif et de langage [2, 5, 11, 12, 13]. Les premiers Sapiens modernes sur le plan comportemental avaient également des comportements plus prosociaux que les hominidés archaïques : ils maintenaient des groupes sociaux plus importants, établissaient des réseaux sociaux réciproques pour le commerce à distance, et coopéraient régulièrement les uns avec les autres dans des groupes composés partiellement ou entièrement de non-apparentés, tout en apportant un soutien altruiste et une coopération aux non-apparentés qui élevaient des enfants ou étaient handicapés [11, 14, 15]. Sur le plan comportemental, les Sapiens modernes se distinguent également par leur longévité saine, comme en témoigne leur résistance au froid et à d’autres conditions climatiques extrêmes [16], leurs besoins énergétiques moindres et la réduction de la mortalité due aux blessures et aux maladies [17, 18, 19], ainsi qu’une durée de vie post-reproductive prolongée qui facilite la reproduction coopérative [11, 20, 21], autant de facteurs qui ont amélioré leur santé et leur viabilité dans des habitats divers, rudes et imprévisibles à travers le monde.
On pense que les lignées de Sapiens et d’Homo neanderthalensis (Néandertaliens) ont divergé à partir d’un ancêtre commun au cours du Pléistocène moyen, avant 500 000 ans (kya), à une époque où la lignée de Sapiens était isolée en Afrique et celle des Néandertaliens était confinée en Europe et en Asie occidentale [22, 23]. On trouve des traces des Néandertaliens datant d’au moins -430,000 en Europe [24], mais les comportements et les génomes des Néandertaliens eux-mêmes sont mieux connus grâce à des artefacts et des fossiles datant de -130,000 à -40,000 ans en Eurasie. En revanche, Sapiens, anatomiquement moderne, est apparu en Afrique de l’Est [22] entre -200,000 et -160,000 ans [2, 25] après une période de -320,000 à -200,000 ans marquée par des fluctuations climatiques imprévisibles [15, 25, 26] qui se sont superposées à une longue période d’aridité progressive [27]. Dans ces conditions écologiques difficiles, les précurseurs des Sapiens modernes sur le plan comportemental ont commencé à entretenir des groupes sociaux plus importants et des alliances sociales réciproques avec des non-apparentés (par exemple, des réseaux commerciaux éloignés), à s’exprimer de manière symbolique (par exemple, l’art, l’ornementation), à collecter des ressources éloignées (par exemple, des pigments, de l’obsidienne et d’autres matériaux), et à faire des recherches sur les ressources naturelles (les pigments, l’obsidienne et d’autres pierres spéciales) pour un usage ultérieur, utiliser des options alimentaires élargies (par exemple, la pêche et la collecte de coquillages) en période de disponibilité imprévisible des ressources, et ont commencé à accumuler des connaissances culturelles et des technologies standardisées qui ont amélioré leur adaptabilité et leur bien-être [15, 25, 26, 28]. Des découvertes récentes suggèrent toutefois que les Sapiens modernes sur le plan comportemental, dotés d’un ensemble de capacités nettement plus imaginatives et flexibles qui n’avaient pas été observées chez les hominidés d’ici ou d’ailleurs, sont apparus en Afrique vers -100,000 ans et se sont répandus sur tout le continent par la suite [29].
Les pressions écologiques et économiques exercées sur les petits groupes de chasseurs mobiles et musclés de la lignée des Néandertaliens en Europe étaient différentes de celles exercées sur la lignée des Sapiens en Afrique de l’Est [30]. Les Néandertaliens et leurs ancêtres européens étaient moins résistants aux extrêmes climatiques, notamment au froid, et leur chasse aux grands animaux terrestres exigeait des dépenses énergétiques quotidiennes élevées [31]. Néanmoins, les Néandertaliens étaient capables de fonctionner efficacement avant d’entrer en compétition avec les Sapiens. Les Néandertaliens ont pu conserver leur dépendance à l’égard de la chasse aux grands animaux terrestres en se déplaçant hors des zones inhospitalières, sans avoir besoin de développer des liens sociaux plus étroits ou des technologies et des ressources plus efficaces et plus variées [17, 22, 28, 31].
Les pratiques novatrices de Sapiens sont mieux documentées après -50,000 ans, lorsqu’elles ont atteint un stade où l’imagination créatrice des humains modernes s’est manifestée de manière évidente [2, 5, 32]. L’épanouissement de la modernité comportementale à la fin du Pléistocène a probablement été facilité par des processus culturels et neurobiologiques progressifs grâce auxquels des comportements complexes, comme l’art figuratif narratif et le langage, sont apparus par exaptation et recrutement comportemental [2]. Quoi qu’il en soit, les caractéristiques de base de la modernité comportementale devaient déjà être présentes lorsque Sapiens s’est répandu hors d’Afrique entre -65,000 et -55,000 ans, alors que le climat africain devenait plus sec et plus froid [27, 33]. Les Sapiens, inventifs, sociables et résilients, ont su s’adapter à des conditions imprévisibles et diverses lors de leur migration hors d’Afrique et se sont répandus dans le monde entier, remplaçant tous les autres hominidés vers -40,000 ans et produisant des cultures qui ont prospéré en continuant à se développer en termes de connaissances, d’art, de science, de technologie et de densité de population jusqu’à aujourd’hui.
Qu’est-ce que la créativité ? Comment la mesure-t-on ?
Le trait le plus distinctif et le plus marquant de la modernité comportementale identifié par les paléoanthropologues et les archéologues est ce que les psychologues ont décrit comme la créativité, en particulier les réalisations et les traits de personnalité des personnes hautement créatives. La créativité peut être définie succinctement comme l’utilisation de l’imagination ou d’idées originales pour atteindre des objectifs importants [34, 35], et c’est un phénomène à multiples facettes qui peut être évalué en fonction de certains aspects de l’intelligence et/ou de certains aspects de la personnalité [36, 37, 38]. Les tests psychométriques des aspects créatifs de l’intelligence ont été développés par Guilford et Torrance pour mesurer les aspects de la pensée divergente dans des tâches verbales et picturales. La pensée divergente est une façon innovante de résoudre des problèmes en explorant de nombreuses solutions possibles, en établissant des liens intuitifs spontanés entre des phénomènes qui sont conventionnellement considérés comme disparates, tout en tolérant une certaine ambiguïté [39]. La pensée divergente se produit généralement dans des états d’évaluation reposante et ludique des pensées et des images internes, tels que le vagabondage de l’esprit en mode par défaut, le flux, l’association libre, la rêverie ou la contemplation [37, 40, 41, 42], qui dépend de l’activation du cortex préfrontal médian pour l’évaluation des stimuli internes en tant que composant central du réseau de conscience de soi [43, 44]. En revanche, la pensée convergente suit une séquence logique d’inférences pour arriver à une solution unique avec certitude ; elle dépend des cortex préfrontal et pariétal latéraux, qui sont des composantes essentielles du réseau exécutif de contrôle de soi qui soutient l’utilisation intentionnelle des symboles et l’inhibition intentionnelle des impulsions déclenchées de l’extérieur [19, 45, 46].
Les caractéristiques utilisées pour mesurer la pensée divergente comprennent l’originalité (pensées inventives et imaginatives), la flexibilité (capacité à passer d’un champ conceptuel à un autre), la fluidité (libre circulation de nombreuses idées et réponses pertinentes), l’élaboration (nombreux détails vifs et spécifiques), un degré élevé d’abstraction et la persistance malgré l’incertitude [39, 47, 48]. Les tests de pensée divergente mis au point par Guilford et Torrance sont les tests les plus utilisés pour évaluer le fonctionnement intellectuel créatif, car ils permettent de prédire fortement les réalisations créatives et la capacité à résoudre des problèmes dans la vie quotidienne [38, 47, 48].
Comment la créativité est-elle liée à d’autres aspects de la modernité comportementale ?
Outre ses propriétés cognitives, la pensée divergente implique des états détendus de conscience intuitive qui se caractérisent également par une spontanéité physique, un affect joyeux, un esprit ludique et une sociabilité [40, 49, 50], qui peuvent être quantifiés en termes de caractéristiques de la personnalité. La personnalité fait référence à la façon dont un individu apprend à façonner et à s’adapter à un environnement interne et externe en constante évolution [51].
Comme la pensée divergente, les caractéristiques de la personnalité créative ont de multiples facettes, y compris des traits de caractère (c’est-à-dire des styles d’autogestion rationnelle, avec des fonctions exécutives d’autodirection, des fonctions législatives de coopération et des fonctions judiciaires d’autotranscendance) et des traits de tempérament (c’est-à-dire des pulsions émotionnelles de curiosité à l’égard de ce qui est nouveau, de volonté de prendre des risques, de volonté de travailler pour la reconnaissance sociale et de persévérance pour l’amour de la réussite) [36, 52, 53]. Les deux domaines du tempérament et du caractère montrent clairement que le potentiel de créativité d’une personne ne peut se développer sans la sagesse de reconnaître ce qui est valable et la plasticité de s’adapter en conséquence. Le test psychométrique le plus largement utilisé pour évaluer les deux domaines de la personnalité créative est l’inventaire du tempérament et du caractère (TCI) [51, 52, 54]. Les tests de pensée créative divergente dans des tâches verbales et picturales, les traits de personnalité créatifs mesurés par le TCI et les évaluations directes des réalisations créatives au cours de la vie sont tous très fiables et validés par leurs fortes corrélations les uns avec les autres, même lorsque l’intelligence générale et les variables démographiques sont contrôlées [47, 48]. De manière empirique, le profil de personnalité créative TCI mesure également la santé humaine en général, y compris le bien-être physique, mental et social [19, 53, 55]. En d’autres termes, les trois domaines de caractéristiques de la modernité comportementale identifiés par les anthropologues sont en fait des aspects interdépendants de la santé et du bien-être de l’homme moderne. Par conséquent, l’ITC fournit des mesures phénotypiques quantitatives valides permettant d’étudier les fonctions cognitives, émotionnelles et sociales, la connectivité cérébrale et la génétique qui sous-tendent la créativité, la prosocialité et d’autres aspects du bien-être chez les humains modernes, et ce, de manière robuste et reproductible [19, 56].
Les spécialistes des sciences cognitives ont proposé que la capacité créative de Sapiens à voir le monde et les autres personnes d’une manière nouvelle dépend de plusieurs processus cérébraux interdépendants d’apprentissage et de mémoire. On pense que la créativité de Sapiens dépend des fonctions cérébrales humaines d’apprentissage prospectif (c’est-à-dire l’encodage, le stockage et la récupération des actions prévues), d’apprentissage constructif (c’est-à-dire le souvenir du passé et l’imagination de l’avenir) et des capacités connexes de théorie de l’esprit (c’est-à-dire la capacité d’attribuer des états mentaux à nous-mêmes et aux autres pour faciliter l’interaction sociale empathique), du mode par défaut (c’est-à-dire la conscience du milieu interne sans se concentrer sur les tâches externes), de la mémoire autobiographique et de la mémoire de la vie quotidienne, (c’est-à-dire la conscience du milieu interne sans se concentrer sur des tâches externes), la mémoire autobiographique (c’est-à-dire le souvenir vivace d’expériences passées avec la conscience contextuelle du moment et du lieu où les faits ont été appris), et la narration d’histoires (c’est-à-dire la composition significative d’art figuratif narratif et de langage), qui sont tous des aspects de la conscience de soi et partagent des circuits cérébraux qui se chevauchent largement [57,58,59]. À leur tour, ces processus sous-jacents à la pensée divergente fonctionnent en coopération avec d’autres processus sous-jacents à la pensée convergente, qui ont des fonctions complémentaires pour la résolution de problèmes dans une vie quotidienne réussie [39, 46]. Tout comme la personnalité créative est multiforme, les fonctions cérébrales qui lui sont associées sont également des aspects multifacettes de systèmes neurocognitifs complexes d’adaptabilité qui sont mesurés par le TCI [19].
Comment les capacités d’adaptation complexes se développent-elles et évoluent-elles ?
Les traits adaptatifs complexes sont organisés par des processus de développement [60] et d’évolution [61,62,63] caractérisés par la multi-finalité (c’est-à-dire que les mêmes antécédents peuvent avoir des résultats différents, comme dans la pléiotropie génotypique) et l’équi-finalité (c’est-à-dire que des antécédents différents peuvent avoir le même résultat, comme dans l’hétérogénéité des chemins génotypiques redondants), comme nous l’avons étudié en détail pour l’architecture génotypique-phénotypique du TCI [19, 64, 65]. Ces propriétés des systèmes complexes sont à la base du rôle important de l’exaptation, c’est-à-dire lorsque des caractères déjà présents sont cooptés pour permettre de nouvelles fonctions adaptatives [66]. Cette plasticité permet à la créativité de faire du neuf avec du vieux ou, plus précisément, de produire des phénotypes et des génotypes adaptatifs complexes par le biais d’interactions dynamiques non linéaires entre les caractéristiques constitutives afin de sélectionner de nouvelles fonctions avantageuses [23]. Le développement de telles fonctions adaptatives complexes est susceptible d’être sélectionné positivement au cours de l’évolution lorsqu’elles sont bénéfiques pour la survie et la reproduction, comme cela a été suggéré pour l’évolution de la créativité chez Sapiens en réponse aux fluctuations climatiques imprévisibles et à la variabilité des ressources qui menaçaient la survie [26], ou lorsque des groupes sociaux et des réseaux commerciaux importants et coopératifs ont commencé à bénéficier d’une communication améliorée par le langage pour faciliter la communication [2, 7, 67].
En raison de l’incomplétude des archives archéologiques, une controverse importante a eu lieu sur la question de savoir si les caractéristiques de la modernité comportementale sont apparues en une seule fois à la fin du Pléistocène [2, 5], ou si certaines caractéristiques sont apparues individuellement et/ou successivement et se sont ensuite organisées par étapes progressives en Afrique en réponse à une pression environnementale accrue après que l’ancêtre commun des Sapiens et des Néandertaliens se soit dispersé en Europe avant -500,000 an [15, 25, 68]. Les informations les plus récentes suggèrent l’adoption de comportements plus complexes après -400,000 et l’émergence des caractéristiques les plus distinctives de la modernité comportementale après -100,000 [25, 26], ce qui suggère le rôle de nombreux gènes dans des réseaux coordonnés, comme prévu pour des traits adaptatifs aussi complexes. Les comportements complexes observés avant et après -100,000 ans se distinguent peut-être mieux par la différence entre la pensée convergente et la pensée divergente, car la meilleure documentation de l’imagination créative moderne est l’art figuratif narratif [2, 5, 32], qui exige à la fois symbolisme et conscience de soi. Les preuves de cette pensée créative divergente apparaissent pour la première fois après -50,000 ans, et les réalisations créatives de Sapiens continuent de s’accumuler jusqu’à aujourd’hui [11].
Quoi qu’il en soit, les caractéristiques de la modernité comportementale sont certainement des traits adaptatifs complexes qui ne peuvent être compris en se concentrant sur une fonction cérébrale ou un gène à la fois. Démêler les complexités de la modernité comportementale présente de nombreux défis de taille.
Horreur
Augmentée
Sélection de textes de
Zero HP Lovecraft
Horreur
Augmentée
Sélection de textes de
Zero HP Lovecraft
Les défis de la compréhension de l’évolution de la créativité humaine
Les comparaisons entre Sapiens et d’autres primates anthropoïdes vivants fournissent des preuves circonstancielles que les changements dans les circuits cérébraux et les fonctions connexes pour le symbolisme et/ou la conscience de soi expliquent les caractéristiques créatives qui distinguent les humains modernes des autres hominidés sur le plan comportemental [12, 45, 57]. Cependant, les fossiles crâniens ne fournissent que des informations limitées sur le cerveau des Néandertaliens et d’autres hominidés disparus [12, 45, 69, 70, 71]. Les preuves archéologiques indiquent que les cultures et les technologies de l’homme de Neandertal ne présentaient guère l’esprit d’innovation qui animait leurs homologues parmi les Sapiens qui les ont remplacés en Europe et en Asie occidentale à partir d’il y a plus de 40,000 ans [9]. Les Néandertaliens avaient des compétences exécutives sophistiquées et produisaient occasionnellement des expressions pouvant être interprétées comme symboliques, mais tous les artefacts qui le suggèrent sont datés d’après 130 kya [17] et surtout d’environ -40,000 après que les Sapiens aient commencé à migrer hors d’Afrique [72,73,74,75]. Il est clair que l’utilisation créative de symboles par les Néandertaliens ne faisait pas partie de la routine de leur vie et de leur culture, et que, bien que sans aucun doute complexe, la relation des Néandertaliens à l’environnement qui les entourait – et sans doute aussi à leurs congénères – était profondément différente de celle qu’entretiennent aujourd’hui les Sapiens [5, 6, 9].
D’autre part, les Néandertaliens étaient de très proches parents de Sapiens, et partageaient sans aucun doute certaines de leurs fonctions comportementales, émotionnelles et cognitives [1,2,3, 12, 45]. Comme l’a exprimé de manière informelle le paléogénéticien Svante Pääbo, “je veux savoir ce qui a changé chez les humains tout à fait modernes, par rapport aux Néandertaliens, et qui a fait la différence. Qu’est-ce qui nous a permis de construire ces énormes sociétés, de nous répandre dans le monde entier et de développer la technologie dont personne ne peut douter qu’elle est unique à l’homme ? Il doit y avoir une base génétique pour cela, et elle se cache quelque part dans ces listes [de paires de bases nucléotidiques des génomes humains] [76].”
Pääbo reconnaît que les progrès réalisés pour répondre à cette question ont été limités pour deux raisons principales : premièrement, un grand nombre de changements dans le génome humain après sa divergence avec l’ancêtre commun des humains et des chimpanzés il y a 7 à 10 millions d’années, et deuxièmement, le manque de connaissance des conséquences fonctionnelles de ces changements [1]. Les progrès ont également été limités par le manque de connaissance de l’architecture génotypique-phénotypique complexe des traits liés à la créativité humaine et à la modernité comportementale : les gènes qui influencent les aspects complexes de la personnalité humaine, tels que la créativité, le symbolisme, la prosocialité et le langage, sont susceptibles d’impliquer de nombreux gènes agissant dans des réseaux coordonnés, plutôt qu’indépendamment [77]. Afin de contourner ces problèmes, nous avons commencé par caractériser les relations génotypiques-phénotypiques complexes qui décrivent l’architecture de la personnalité humaine moderne en utilisant le TCI [19, 64, 65].
Les relations génotype-phénotype qui sous-tendent la modernité comportementale
Nous avons évalué la personnalité humaine moderne à l’aide de l’ITC parce qu’il fournit des mesures hautement fiables et empiriquement validées de la créativité et d’autres aspects de la modernité comportementale qui sont héréditaires et fondés sur la neurobiologie, y compris les aspects physiques, émotionnels, sociaux, cognitifs et spirituels du bien-être [19, 42, 47, 56, 78, 79] aussi bien, voire mieux, que d’autres tests disponibles [54]. Le TCI tient compte de deux domaines de la personnalité basés sur des formes distinctes d’apprentissage et de mémoire : le tempérament (c’est-à-dire la composante inconsciente de la personnalité – les habitudes conditionnées par l’association et la réactivité émotionnelle) et le caractère (c’est-à-dire les composantes autorégulatrices de la personnalité – ce que les gens font d’eux-mêmes de manière intentionnelle et/ou créative) [51]. Il a été développé comme une mesure complète de la personnalité humaine, et capture les caractéristiques de la modernité comportementale, y compris la créativité et le comportement prosocial, comme indiqué dans la section précédente et ailleurs [19, 42, 47, 48, 54, 56, 78, 79]. Les indices TCI de créativité et de bien-être permettent également de prédire les mesures subjectives et objectives de la santé physique, y compris la longévité en bonne santé [19, 55, 80].
Nous avons utilisé des méthodes axées sur les données pour mener des études d’association à l’échelle du génome de l’ITC dans trois échantillons différents ayant des environnements et des cultures différents (Finlandais, Allemands et Coréens). Nous avons ainsi pu déconstruire les réseaux génotypiques-phénotypiques complexes et les interactions environnementales qui sous-tendent le tempérament et le caractère de l’homme moderne [64, 65]. Ces méthodes tiennent correctement compte des propriétés des systèmes adaptatifs complexes, notamment la pléiotropie et l’hétérogénéité génétique [81].
Plus précisément, nous avons procédé par étapes pour caractériser les relations phénotypiques-génotypiques à plusieurs niveaux d’organisation. Tout d’abord, dans un échantillon épidémiologiquement représentatif de 2149 Finlandais, nous avons identifié des ensembles de polymorphismes mononucléotidiques (SNP) qui se regroupent naturellement chez des individus particuliers, indépendamment du phénotype. Deuxièmement, nous avons découvert cinq groupes de personnes présentant des configurations distinctes des 13 facettes du domaine d’autorégulation de la personnalité humaine (c’est-à-dire les dimensions de caractère de l’autodirection, de la coopération et de l’autotranscendance), indépendamment du génotype. Troisièmement, nous avons trouvé 42 ensembles de SNP qui étaient significativement associés aux profils de caractère et identifié 727 loci génétiques. Nous avons reproduit 95 % des 42 ensembles SNP dans un échantillon de 902 Allemands en bonne santé et un échantillon de 1092 Coréens, ainsi que leur association avec les groupes de caractères [64]. Les ensembles génotypiques associés au caractère se sont avérés moduler des processus moléculaires spécifiques dans le cerveau pour la fixation intentionnelle d’objectifs, l’autoréflexion, l’empathie, l’apprentissage et la mémoire épisodiques, et la longévité en bonne santé.
Il est remarquable de constater que 68 % des 727 gènes associés aux caractères étaient uniques à un seul profil de caractère [64]. Par conséquent, de multiples groupes de gènes ont conduit à chaque trait de caractère individuel. Par exemple, un haut degré d’autonomie se produit chez différents individus par le biais de processus moléculaires distincts des réseaux génotypiques qui dépendent de configurations particulières de l’autonomie avec d’autres aspects des traits de caractère. En d’autres termes, les gènes de l’autonomie étaient différents chez les personnes présentant le profil de caractère créatif (les trois dimensions du caractère sont élevées, de sorte que les objectifs valorisés sont désintéressés, prosociaux et altruistes), le profil organisé (l’autonomie et la coopération sont élevées, mais le dépassement de soi est faible, ce qui permet une action répondant à la fois aux besoins personnels et sociaux pour un bénéfice mutuel, mais sans sacrifice pour les autres), ou le profil ingénieux (seule l’autonomie est élevée, ce qui entraîne des motifs égocentriques).
Ensuite, nous avons répété ce processus avec les 12 facettes des pulsions émotionnelles humaines (c’est-à-dire les dimensions du tempérament que sont la recherche de la nouveauté, l’évitement du mal, la dépendance à la récompense et la persistance) [65]. Nous avons mis en évidence trois groupes de personnes présentant des profils de tempérament distincts, indépendamment du génotype. L’un de ces groupes était caractérisé par une faible recherche de nouveauté, une forte dépendance à la récompense et une forte persistance, que nous avons désigné comme l’ensemble de tempéraments fiables, comme nous l’avons expliqué en détail ailleurs [56, 82]. Les autres groupes de tempérament étaient le groupe antisocial et le groupe d’hypersensibilité émotionnelle. 51 ensembles de SNP ont été associés de manière significative aux groupes de tempérament. Les 736 gènes associés à ces ensembles de SNP étaient enrichis dans les voies moléculaires activées par le conditionnement associatif chez les animaux, y compris les voies ERK, PI3K et PKC qui sont cruciales pour la modulation de la plasticité synaptique et l’apprentissage à long terme impliqués dans le conditionnement associatif de la réactivité émotionnelle, de l’attachement social et de la persistance. Nous avons reproduit 90 % des 51 ensembles de SNP pour les groupes de tempérament dans les échantillons allemands et coréens sains.
Les gènes que nous avons découverts pour le tempérament et le caractère se chevauchaient partiellement, nous avons donc évalué l’organisation des groupes de tempérament et de caractère conjointement. Nous avons découvert trois réseaux phénotypiques qui expliquent les relations conjointes des groupes de traits de tempérament avec les groupes de traits de caractère. Nous avons désigné ces groupes conjoints tempérament-caractère comme étant (i) le réseau émotionnel peu fiable (c.-à-d., les personnes très réactives sur le plan émotionnel avec peu de maîtrise de soi ou de créativité), (ii) le réseau organisé-fiable (c.-à-d., les personnes ayant une forte maîtrise de soi, des conflits et des objectifs émotionnels, mais peu de créativité), et (iii) le réseau créatif-fiable (c.-à-d., les personnes très créatives, prosociales et perspicaces dans leur évaluation des valeurs et des théories) [19]. Nous avons constaté que ces réseaux phénotypiques étaient presque disjoints (c’est-à-dire qu’ils partageaient peu de sujets ou de caractéristiques phénotypiques) (voir figure supplémentaire 1). Chacun des trois réseaux phénotypiques était fortement corrélé avec un réseau génotypique multi-locus différent (voir les figures supplémentaires S2 et S3).
Les fonctions des gènes qui ont été mis en correspondance avec les réseaux génotypiques se sont avérées réguler des systèmes distincts d’apprentissage et de mémoire qui sous-tendent la personnalité : (i) un réseau multi-locus de 249 gènes pour la régulation de la réactivité émotionnelle, du conditionnement associatif et des attachements sociaux, que nous avons désigné comme le réseau de la “réactivité émotionnelle” ; (ii) un réseau multi-locus de 438 gènes pour la régulation de la recherche d’objectifs intentionnels, tels que l’acquisition volontaire de nourriture, la fabrication d’outils, le travail d’équipe coopératif, l’analyse logique et la symbolisation, que nous avons désigné comme le réseau du “contrôle de soi” ; et (iii) un réseau génotypique de 574 gènes pour l’apprentissage épisodique et la mémoire autobiographique de la vie d’une personne en tant que récit du passé, du présent et du futur dans lequel la personne peut explorer des perspectives alternatives avec une perspicacité intuitive et une imagination créative, que nous avons désigné comme le réseau “conscience de soi”. Il est remarquable que 73% des 972 gènes de ces trois réseaux soient uniques à un seul réseau. Il est rare de trouver une séparation aussi forte des clusters spécifiés par des ensembles aussi complexes de variables phénotypiques et génotypiques [19].
Les gènes que nous avons identifiés pour le tempérament et le caractère représentaient presque toute l’héritabilité de la personnalité attendue des études de jumeaux [64, 65]. Les fortes relations entre les trois réseaux phénotypiques du tempérament et du caractère et les trois principaux réseaux génotypiques de l’adaptabilité humaine nous ont fourni des outils précieux pour évaluer l’évolution de la créativité humaine et d’autres aspects de la modernité comportementale en comparant les génomes des chimpanzés (Pan troglodytes) et des Néandertaliens à ceux des humains modernes.
Hypothèses à tester
Nous avons émis l’hypothèse que les trois réseaux génotypiques presque disjoints pour l’adaptabilité humaine ont évolué par étapes successives au cours de l’évolution de la personnalité humaine moderne. Pour tester cette hypothèse, nous avons étudié les 972 gènes identifiés pour la personnalité chez Sapiens, dont beaucoup ont également été trouvés dans les génomes des Néandertaliens et des chimpanzés. Bien qu’elles ne soient que très éloignées les unes des autres, ces deux espèces sont les plus proches de l’homme moderne et possèdent des génomes bien caractérisés comparables aux génomes à forte couverture de l’homme moderne [1]. Nous avons émis l’hypothèse que les trois réseaux diffèrent les uns des autres chez Sapiens, Neandertal et les chimpanzés. Plus précisément, nous avons supposé que (i) les chimpanzés auraient des gènes uniquement dans le réseau de la réactivité émotionnelle, (ii) les Néandertaliens et les Sapiens partageraient de nombreux gènes pour l’autocontrôle intentionnel, ce qui était déjà évident dans leur lignée humaine commune, et (iii) les gènes trouvés uniquement chez Sapiens seraient plus fréquents dans le réseau de la conscience créative de soi, comme prédit précédemment sur la base des changements coïncidents dans le cerveau et le comportement au cours de l’évolution des hominoïdes [12, 45]. Une fois que nous avons identifié les gènes uniques à l’homme moderne à partir de ces analyses, nous avons évalué quels types de gènes distinguaient les trois réseaux. Nous avons également évalué les modèles de transmission alternatifs et les conditions environnementales qui pourraient expliquer l’émergence soudaine de la créativité chez l’homme moderne. Enfin, nous avons examiné où s’expriment dans le cerveau les gènes de l’apprentissage et de la personnalité propres à l’homme moderne.
Sujets et méthodes
Sujets humains modernes
Notre échantillon de Sapiens était l’étude Young Finns, une étude épidémiologique de 2149 sujets finlandais en bonne santé qui ont été évalués en 1997, 2001, 2007 et 2012 [83]. Tous les sujets ont fait l’objet d’évaluations génotypiques, environnementales et phénotypiques complètes et standardisées, y compris l’administration du TCI [64, 65].
Adaptation de la nouvelle de
Lovecraft en animation grâce
à l’IA
Adaptation de la nouvelle de
Lovecraft en animation
Indicateurs de personnalité de la modernité comportementale, de la créativité et du bien-être
Les sujets finlandais ont rempli le TCI avec 240 critères auto-rapportés en utilisant une échelle de Likert en 5 points [84]. La consistance interne des échelles et leur fiabilité au test étaient fortes : r > 0,8 pour les dimensions et > 0,65 pour les sous-échelles individuelles entre les suivis, y compris le suivi à 15 ans. Les moyennes des scores des échelles et sous-échelles sur les quatre occasions d’évaluation ont été utilisées pour réduire l’erreur de mesure.
Tous les sujets ont rempli le TCI afin d’évaluer quatre dimensions du tempérament (évitement du mal, recherche de la nouveauté, dépendance à la récompense et persistance) et trois dimensions du caractère (autonomie, coopération et dépassement de soi). Chacune de ces dimensions comporte de multiples facettes (sous-échelles) mesurant l’expression de cette dimension dans différentes situations. Les descriptions des personnes ayant obtenu des scores élevés et faibles pour chaque dimension et ses sous-échelles sont présentées dans le tableau supplémentaire S1.
Nos analyses antérieures de l’architecture génotypique-phénotypique de l’ICC ont révélé une structure hiérarchique naturelle qui est importante pour comprendre les relations complexes des indicateurs phénotypiques des prédispositions génotypiques à la modernité comportementale humaine qui peuvent être dérivés de l’ICC, comme la créativité, le bien-être, la conscience de soi et la maîtrise de soi, comme nous l’avons décrit en détail ailleurs [19, 82] et dans les informations supplémentaires. La personnalité humaine peut être décrite à trois niveaux de complexité à partir (1) des dimensions individuelles du tempérament et du caractère, chacune étant composée de la somme de leurs sous-échelles, comme le montre le tableau supplémentaire S1, (2) des profils de tempérament multi-traits génétiquement indépendants ou des profils de caractère multi-traits [64, 65], et (3) des réseaux conjoints de profils de tempérament et de caractère qui indiquent l’intégration de multiples processus d’apprentissage de chacun des trois principaux systèmes d’apprentissage et d’adaptabilité humains, comme décrit ailleurs [19] et brièvement dans l’introduction.
Nous avons utilisé deux indices de santé dérivés du TCI, un indice de bien-être et un indice de résilience au mal-être. Nous avons confirmé la validité du profil de personnalité créative dans lequel les trois traits de caractère de l’ICC (c’est-à-dire l’autodirection, la coopération et l’autotranscendance) sont hautement développés en tant qu’indice de bien-être dans de multiples échantillons d’humains modernes de différentes cultures [19, 52, 55, 80]. De même, si l’on ne tient pas compte de l’autotranscendance, la somme de l’autodirection et de la coopérativité est un indicateur de la résilience face au mal-être dans de nombreuses cultures [19, 52, 55, 80]. Dans notre échantillon de Finlandais, nous avons confirmé la validité de ces deux indices à l’aide de mesures indépendantes de l’équilibre affectif positif, du soutien social perçu, des comportements physiques (exercice, tabagisme, alimentation) et des résultats de laboratoire objectifs pour une santé idéale tels que recommandés par l’American Heart Association, comme décrit ailleurs [19] et dans le tableau supplémentaire S2.
Annotation des gènes
L’étude a été réalisée avec 972 gènes cartographiés sur les trois réseaux phénotypiques : Créatif-fiable, Organisé-fiable et Émotionnel-non fiable (figure supplémentaire S1) [19]. Nous appelons les réseaux génotypiques correspondants les réseaux de conscience de soi, de maîtrise de soi et de réactivité émotionnelle, respectivement (tableau supplémentaire S1 et figures S2 et S3). Les annotations des gènes individuels ont été obtenues à l’aide de l’API perl d’Ensembl [85] versions 87-92 (tableau supplémentaire S3) et classées selon leur biotype en distinguant les gènes codant pour des protéines, les gènes à ARN non codant et les pseudogènes (tableau supplémentaire S4).
Génomique comparative
Les orthologues du chimpanzé pour les 972 gènes liés à la personnalité chez l’Homo sapiens moderne ont été obtenus en accédant à la base de données CHIMP2.1.4, qui utilise le modèle Pan troglodytes (7/20/16) construit à partir du génome (v.2.1.4) avec des fichiers de modèles de gènes (R.89) d’Ensembl en utilisant l’API Perl [86]. Les gènes orthologues d’autres primates (Bonobo, Chimpanzé, Gibbon, Gorille, Humain, Macaque, Oursin et Orang-outan) ont été obtenus en utilisant l’accès programmatique aux ressources d’Ensembl [87].
Les orthologues néandertaliens des 972 gènes liés à la personnalité chez Sapiens ont été identifiés dans les données annotées du Neanderthal Genome Project [88]. La reproductibilité de ces résultats a ensuite été évaluée dans des analyses séparées des génomes à haute couverture du Néandertalien de l’Altaï [88], d’un autre Néandertalien de la grotte Vindija [89] (spécimen 33.19, http://cdna.eva.mpg.de/neandertal/Vindija), et d’un troisième de la grotte Chagyrskaya 90. Ces analyses nous ont permis d’identifier les gènes que les chimpanzés et/ou les Néandertaliens partageaient avec les humains modernes parmi ceux qui n’étaient présents que chez ces derniers (tableau supplémentaire S5), puis de comparer leurs caractéristiques (tableaux supplémentaires S6 et S7).
Méthodes statistiques générales
Nous avons utilisé l’analyse de la variance (ANOVA) pour tester l’hypothèse nulle selon laquelle les trois réseaux étudiés sont similaires en termes de gènes qui les composent au sein d’une même espèce et entre espèces (Sapiens, Chimpanzé et Neandertal). Pour ce faire, nous avons utilisé l’ANOVA pour les échantillons indépendants et corrélés, un par réseau (Conscience de soi vs Contrôle de soi vs Réactivité émotionnelle) dans chacune des espèces. Nous avons ensuite appliqué des comparaisons post-ANOVA à l’aide du test des différences les plus significatives (HSD) de Tukey pour évaluer les différences spécifiques entre les paires de réseaux (par exemple, Créatif vs Organisé). Nous avons utilisé le test ANOVA tel qu’implémenté dans Concepts & Applications of Inferential Statistics, Richard Lowry 1998-2021, http://vassarstats.net/anova1u.html, et dans le paquet rstatix de R.
Pour plus de clarté dans la présentation des résultats, les valeurs p indiquées dans les descriptifs des tests ANOVA et Tukey’s HSD ont été arrondies aux valeurs de signification plus conservatrices de 0,0001 et 0,01, respectivement, qui ont effectivement tendance à dépasser les valeurs E-30 et E-10, respectivement. La taille de l’effet de l’ANOVA a été calculée comme la valeur f définie par Cohen [91], qui propose l’interprétation suivante de cette valeur : f = 0,1 est un petit effet, f = 0,25 est un effet moyen et f = 0,4 est un grand effet. Tous les autres paramètres utilisés dans chaque mesure de l’ANOVA ont été calculés comme d’habitude [91, 92], et les résumés complets de toutes nos ANOVA sont fournis dans les informations supplémentaires et les tableaux supplémentaires S8-S12.
Estimation génotypique de la modernité comportementale des Néandertaliens
Le nombre de gènes individuels que les Néandertaliens partageaient avec les humains modernes n’est peut-être pas un indicateur adéquat de leur impact sur la créativité et d’autres aspects du fonctionnement de l’homme moderne. Nous avons donc évalué l’impact des gènes sur la prédisposition au bien-être de l’homme moderne comme indicateur de la modernité comportementale en estimant leurs rôles relatifs dans des ensembles de SNP spécifiques afin de prendre en compte les interactions entre les ensembles coordonnés de gènes qui ont un impact sur le bien-être.
Afin d’extraire des échantillons prototypiques d’humains présentant des caractéristiques distinctives de type néandertalien et des caractéristiques distinctives de type sapiens, nous avons d’abord identifié les gènes présents uniquement chez Sapiens et les gènes que Neandertal partageait avec Sapiens, à l’exclusion des gènes présents chez les chimpanzés (Tableau supplémentaire S3). Nous avons ensuite effectué une corrélation croisée entre ces gènes et les ensembles de SNP originaux dans lesquels ils avaient été détectés en relation avec le caractère [64] et/ou le tempérament [65] (voir Informations supplémentaires). Nous avons sélectionné des ensembles de SNP trouvés dans les réseaux génotypiques pour la conscience de soi, le contrôle de soi et la réactivité émotionnelle, pour lesquels nous avions déjà mesuré les niveaux de fonctionnement associés chez les humains modernes, y compris deux indices (bien-être et résilience au mal-être) [19]. À partir des mesures de bien-être dont nous disposions pour les ensembles de SNP qui contenaient un ou plusieurs des gènes que les Néandertaliens partageaient avec les Sapiens, nous avons ensuite estimé le bien-être moyen des humains semblables aux Néandertaliens en pondérant le bien-être des personnes dans ces ensembles de SNP individuels par la proportion de gènes présents chez les Néandertaliens par rapport aux Sapiens dans cet ensemble de SNP pour chacun des réseaux. De même, nous avons estimé le bien-être moyen des humains prototypiques de type Sapiens à partir des mesures de bien-être dans les ensembles de SNP qui contenaient un ou plusieurs des gènes présents uniquement chez les humains modernes. Enfin, nous avons comparé les niveaux de bien-être pondéré dans les ensembles de SNP des humains de type Néandertal et des prototypes humains de type Sapiens à l’aide de statistiques ANOVA, y compris les comparaisons post-ANOVA et les tailles d’effet. Enfin, nous avons estimé la modernité génotypique relative de ces prototypes pour les deux espèces à partir du rapport de leurs niveaux moyens de bien-être.
Transfert horizontal de gènes (THG)
Afin de déterminer si les gènes cartographiés dans les trois réseaux phénotypiques ont pu être acquis horizontalement, nous avons calculé leur chevauchement avec les régions de HGT identifiées par Huang et al [88] dans le génome de référence humain hg 19 [93].
Fréquence allélique dérivée (DAF)
Nous avons comparé les scores DAF pour les gènes d’ARN à longue durée de vie et non codants (linc) (tableau supplémentaire S4) trouvés chez les Néandertaliens avec ceux trouvés uniquement chez les Sapiens afin de tester la sélection différentielle (tableaux supplémentaires S6 et S7). Les scores DAF [94] ont été calculés pour les gènes de lincRNA, y compris leurs exons et promoteurs, à l’aide de l’outil AnnLoc (http://annolnc.cbi.pku.edu.cn). Les scores DAF de 0,1 ou moins sont associés à une diversité réduite indiquant une sélection purificatrice (c’est-à-dire une sélection négative contre les allèles délétères), tandis que les scores DAF >0,1 indiquent une diversité accrue, comme cela peut se produire avec des allèles non fonctionnels, une sélection positive d’allèles avantageux ou l’ajout de nouveaux allèles avantageux dans les gènes par HGT [95, 96].
Co-expression des gènes dans le cerveau
Pour évaluer davantage les fonctions des gènes que nous avons trouvés uniquement chez Sapiens, nous avons utilisé Process Genes List pour analyser les listes de gènes qui correspondent à des ensembles de SNP particuliers avec au moins un gène trouvé uniquement chez Sapiens [97]. Cette méthode d’apprentissage automatique utilise le Allen Brain Atlas pour calculer un niveau d’expression moyen normalisé de l’ARNm dans chaque région du cerveau pour les listes de chaque ensemble de gènes. Les régions du cerveau dans lesquelles ces gènes étaient significativement co-exprimés ont été identifiées et affichées dans les images du cerveau.
De plus amples détails sur toutes nos méthodes et analyses statistiques sont disponibles en tant qu’informations supplémentaires.
Résultats
Les réseaux génotypiques de personnalité distinguent Sapiens des autres hominoïdes
Nous avons d’abord testé lesquels des 972 gènes associés aux réseaux conjoints tempérament-caractère de Sapiens (tableau supplémentaire S3) étaient également présents dans les génomes des Néandertaliens et/ou des chimpanzés. Nous avons trouvé 509 gènes de la personnalité humaine moderne chez les trois hominoïdes, 148 chez les Néandertaliens mais pas chez les chimpanzés, 48 chez les chimpanzés et pas chez les Néandertaliens, et 267 uniquement chez Sapiens (Tableau 1).Tableau 1 Analyse statistique des 972 gènes de la personnalité humaine moderne par réseau de personnalité et par espèce : Tableau 1 Analyse statistique des 972 gènes de la personnalité humaine moderne par réseau de personnalité et par espèce : Néandertaliens (N) et chimpanzés (C)*.
Nous avons émis l’hypothèse que les gènes correspondant aux réseaux génotypiques de la réactivité émotionnelle, du contrôle de soi et de la conscience de soi seraient présents de manière différentielle dans les génomes des chimpanzés, des Néandertaliens et des Sapiens. Pour vérifier cela, nous avons effectué une ANOVA pour les gènes des trois réseaux ainsi que des contrastes des paires d’espèces possibles en supposant des échantillons corrélés (comme cela se produit avec l’héritage vertical des parents à la progéniture) ou des échantillons indépendants (comme cela se produit avec le HGT d’organismes autres que les parents), comme résumé dans le tableau 2 et décrit dans le tableau supplémentaire S8. Nous avons constaté que les trois espèces diffèrent significativement les unes des autres pour chacun des trois réseaux, que les échantillons soient corrélés ou non (239 > F (2[1664,2913]) > 16, p < 0,0001) (Tableau 2, Fig. 1, Tableau supplémentaire S8). Les différences entre les espèces étaient importantes pour les gènes liés à la conscience de soi (taille de l’effet de Cohen f = 0,46), intermédiaires pour la maîtrise de soi (f = 0,35) et faibles pour la réactivité émotionnelle (f = 0,21) (tableau supplémentaire S8).Tableau 2 Analyse de variance à sens unique (ANOVA, p < 0. 0001) pour la signification des différences du nombre des 972 gènes associés à la personnalité chez l’homme moderne entre les trois espèces (ANOVA, p < 0,0001) avec les contrastes des nombres de ces gènes dans les paires des espèces selon que les échantillons sont supposés être corrélés ou indépendants. La signification de chaque comparaison corrigée pour le nombre de tests est indiquée. Les espèces comprennent l’Homo sapiens moderne (“Sapiens”), l’Homo neanderthalensis (“Neanderthals”) et Pan troglodytes (Chimpanzés).Tableau complet
Dans les comparaisons par paire, Sapiens diffère significativement des Néandertaliens ainsi que des chimpanzés en ce qui concerne les gènes présents dans chacun des trois réseaux, que les échantillons soient corrélés ou non (test HSD de Tukey, p < 0,01) (tableau 2, tableau supplémentaire S8). Il n’y avait qu’une petite différence entre les Néandertaliens et les chimpanzés en ce qui concerne les gènes présents dans chacun des trois réseaux ; la différence était faiblement significative pour chacun des réseaux si les échantillons sont considérés comme corrélés (test HSD de Tukey, p < 0,05) et non significative pour le réseau de réactivité émotionnelle si les échantillons sont indépendants (tableau 2, tableau supplémentaire S8). Nous examinerons plus loin la possibilité d’un transfert vertical et horizontal dans l’évolution de la personnalité humaine, mais les différences observées ici dans le tableau 2 étaient à peu près les mêmes, que les échantillons soient considérés comme corrélés ou non ; la seule exception était que la plus petite différence (c’est-à-dire entre les Néandertaliens et les chimpanzés pour les gènes régulant la réactivité émotionnelle) n’était pas significative si les échantillons étaient considérés comme indépendants. En résumé, nous avons constaté que les gènes présents dans chacun des trois réseaux génotypiques diffèrent entre chaque paire des trois espèces d’hominoïdes (test HSD de Tukey, p < 0,01 à p < 0,05).
Ensuite, nous avons observé que les gènes des trois réseaux sont représentés chez les hominoïdes sous forme d’ajouts cumulatifs, ce qui est cohérent avec l’hypothèse selon laquelle les Néandertaliens étaient similaires aux chimpanzés en ce qui concerne les gènes de la réactivité émotionnelle, mais étaient intermédiaires entre les chimpanzés et les Sapiens pour ce qui est du nombre de gènes présents dans les réseaux de contrôle de soi et de conscience de soi (Fig. 1). Sur les 972 gènes significativement associés à la personnalité chez Sapiens, 653 étaient présents chez Neandertal et 557 chez les chimpanzés (tableau 1).
Comparés aux chimpanzés, les Néandertaliens ne diffèrent pas dans leurs proportions de gènes de réactivité émotionnelle, que l’on prenne en compte tous les gènes liés à la personnalité dans chaque espèce (tableau 1, par exemple, 71% de 653, 62% de 557), ou que l’on prenne en compte l’ensemble des gènes de la personnalité humaine (18% vs 16% de 972, F (1, 1972) = 1,76, non significatif) (tableau supplémentaire S9). Si l’on met de côté les 54 gènes trouvés uniquement chez l’homme moderne, 72 % des 195 gènes de la réactivité émotionnelle étaient communs aux trois espèces.
Cependant, par rapport aux chimpanzés, les Néandertaliens avaient une plus grande proportion de gènes pour la maîtrise de soi (32 % contre 25 % de 972, F (1, 1942) = 6,86, p < 0,008) et pour la conscience de soi (38 % contre 33 % de 972, F (1, 1942) = 9,1, p < 0,0001) lorsque tous les gènes de la personnalité humaine étaient pris en compte (tableau supplémentaire S9).
En mettant de côté les gènes de la personnalité humaine présents chez les chimpanzés, nous avons constaté que les Néandertaliens n’avaient que 33% des gènes du réseau de la conscience de soi de Sapiens (soit 85 gènes sur 254), 37% des gènes du réseau du contrôle de soi (soit 68 sur 186), 41% des gènes du réseau de la réactivité émotionnelle (soit 38 sur 92). En d’autres termes, si l’on exclut les gènes présents chez les chimpanzés, 67% des gènes de la conscience de soi, 63% des gènes du contrôle de soi et 59% des gènes de la réactivité émotionnelle ne se trouvent que chez Sapiens.
Néanmoins, nous avons reconnu que le nombre de gènes que les Néandertaliens partagent avec les Sapiens pourrait ne pas être un indicateur direct de leur impact sur la créativité et d’autres aspects du bien-être en raison de l’organisation modulaire des gènes dans les systèmes complexes. Plus précisément, nous devions évaluer l’impact des gènes individuels sur le bien-être en estimant leurs rôles relatifs dans des ensembles de SNP spécifiques afin de prendre en compte les interactions entre les ensembles coordonnés de gènes qui ont un impact sur le bien-être.
Nous avons donc estimé l’impact sur le bien-être des gènes partagés par Neandertal et Sapiens par rapport à l’impact sur le bien-être des gènes trouvés uniquement chez Sapiens. Afin d’extraire des échantillons prototypiques d’humains présentant des caractéristiques distinctives de type Néandertalien et des caractéristiques distinctives de type Sapiens, nous avons d’abord identifié les 267 gènes présents uniquement chez Sapiens et les 148 gènes que les Néandertaliens partageaient avec Sapiens, à l’exclusion des gènes présents chez les chimpanzés (Tableau 1, Tableau supplémentaire S3). Nous avons estimé le niveau moyen de bien-être des humains de type néandertalien à partir du bien-être des individus dans les groupes de gènes naturels (c’est-à-dire les ensembles de SNP) qui comprenaient un ou plusieurs des 148 gènes distinctifs des Néandertaliens. Nous avons estimé le niveau moyen de bien-être des humains prototypiques de type Sapiens à partir du bien-être des individus dans des groupes de gènes naturels (c’est-à-dire des ensembles de SNP) comprenant un ou plusieurs gènes trouvés uniquement chez Sapiens.
Le Moment
Straussien
Ebook offert à nos tipeurs
sans montant minimum
Le Moment
Straussien
Ebook offert à nos tipeurs
sans montant minimum
Nous avons constaté que les groupes de gènes de type néandertalien amélioraient le bien-être plus que ce à quoi on s’attendait compte tenu du nombre de gènes qu’ils partageaient avec les humains modernes, mais les niveaux moyens de bien-être étaient toujours systématiquement inférieurs pour les humains de type néandertalien que pour les humains de type sapiens (tableau supplémentaire S10). Les différences entre ces groupes prototypiques étaient similaires dans les comparaisons basées sur deux indices de fonctionnement sain (bien-être et résilience au mal-être) pour chacun des trois réseaux génotypiques, notamment la conscience de soi et la maîtrise de soi (F (3,252) = 34, F (3,454) = 35, p < 0,0001) et la réactivité émotionnelle (F (3, 112) = 15,5, p < 0,0005) (tableau supplémentaire S10). Plus précisément, l’impact des gènes de la conscience de soi sur le bien-être des humains de type Néandertal était de 70 % de celui des Sapiens, ce qui représente une différence assez importante (taille de l’effet f = 0,34). De même, l’impact combiné des gènes de la maîtrise de soi et de la conscience de soi sur le bien-être des humains de type Néandertal était de 67 % de celui de Sapiens (taille de l’effet f = 0,28). L’impact des gènes de la réactivité émotionnelle sur le bien-être des humains de type Néandertal était de 61 % de celui des Sapiens (taille de l’effet f = 0,20). Des résultats similaires ont été obtenus, indiquant une moindre résilience au mal-être chez les individus de type Néandertal (tableau supplémentaire S10).
Les types de gènes distinguent les réseaux de personnalité et les espèces hominoïdes
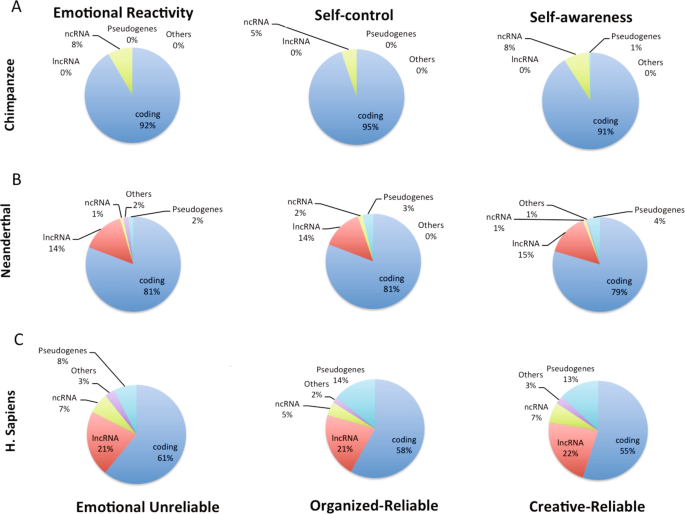
Nous avons analysé les types de gènes pour les réseaux tempérament-caractère de l’homme moderne qui sont présents dans chaque groupe d’hominoïdes, comme le montre la figure 1. Sur les 557 gènes présents chez les chimpanzés, 92 % étaient codants pour les protéines et aucun n’était un ARN long-non codant (lnc) ou un pseudogène. Sur les 653 gènes présents chez les Néandertaliens, 81 % étaient codants pour les protéines, le reste étant des ARNlc (14 %), des pseudogènes (2 %) et des ARN non codants (1 %). Sur les 972 gènes associés à la personnalité chez Sapiens, seuls 61 % étaient codants pour les protéines et il y avait beaucoup de lncRNA (21 %) et de pseudogènes (8 %). Ces distributions variaient peu entre les réseaux au sein de chaque groupe d’hominoïdes, comme prévu par notre hypothèse selon laquelle il y a eu des étapes successives entre les espèces pour améliorer la régulation de l’expression coordonnée de groupes de gènes au sein de chaque espèce (Fig. 1).
La présence de lncRNA et de pseudogènes distingue fortement les types de gènes trouvés dans les trois réseaux de personnalité des humains (Néandertaliens et Sapiens) de ceux trouvés chez les chimpanzés (F (5,1926) = 91,1, p < 0,0001, taille de l’effet f = 0,69, et test HSD de Tukey, p < 1E-23, Fig. 1 et Tableau supplémentaire S11). Les chimpanzés ne présentaient aucun des lncRNA associés à la personnalité humaine moderne (Fig. 1). Les Sapiens avaient plus de lncRNA que les Néandertaliens (21-22 % contre 14-15 %, test HSD de Tukey, p < 1E-12) (Fig. 1). Les différences de gènes régulateurs candidats entre les groupes d’hominoïdes ont été confirmées avec les gènes présents uniquement chez Sapiens (Fig. 2).
Type distinct d’évolution du génome et direction de la sélection chez Sapiens
Nous avons testé la présence de gènes ancestraux impliqués dans la personnalité humaine en recherchant des orthologues dans 57 organismes appartenant aux groupes taxonomiques suivants : Primates, autres Mammifères, Marsupiaux, Monotrèmes, Avians, Reptiles, Amphibiens, Poissons, Cyclostomes, Tuniciers, Insectes, et Nématodes (Tableau supplémentaire S1, Fig. supplémentaire S4A). Nous avons constaté que 557 des 972 gènes liés à la personnalité ont des orthologues dans ces espèces, ce qui suggère une transmission par des ancêtres communs. Les 415 gènes restants, apparemment sans orthologues connus, pourraient avoir été acquis indépendamment. Une transmission indépendante pourrait avoir eu lieu, par exemple, à la suite d’une HGT, qui est largement impliquée dans le génome humain, en particulier chez les primates [88]. Nous avons constaté que 39 gènes associés à la personnalité humaine se trouvaient dans des régions HGT déjà connues (tableau supplémentaire S3). Ainsi, les gènes liés à la personnalité sont enrichis dans les régions HGT connues (4,0% de 972) par rapport au taux global dans le génome humain (1,1%, 642 de 57 905) [88].
Environ 65% des 415 gènes sans orthologues appartiennent au réseau de la conscience de soi, qui est fortement associé au profil de personnalité créative-fiable. En revanche, nous avons constaté que les gènes liés à la personnalité situés dans les régions HGT étaient enrichis dans les trois réseaux génotypiques, avec des taux légèrement plus élevés pour les gènes du réseau de contrôle de soi (5,5 %, 24 gènes sur 438) et du réseau de réactivité émotionnelle (4,0 %, 10 gènes sur 249) que pour le réseau de conscience de soi (3,0 %, 17 gènes sur 574). Seuls deux des gènes dans les régions HGT n’ont été trouvés que chez Sapiens, ce qui suggère que le HGT a eu peu de rôle dans l’émergence de la modernité comportementale.
En revanche, nos découvertes sur les lincRNA ont fourni des preuves soutenant un rôle majeur dans l’émergence de la modernité comportementale. Parmi les 972 gènes associés à la personnalité chez Sapiens, nous avons constaté que 127 étaient des lincRNA sans orthologues dans 57 espèces (voir Informations supplémentaires) : 68 étaient présents uniquement chez Sapiens, 59 chez Neandertal et aucun chez les chimpanzés (Tableau supplémentaire S2). Des informations sur les scores DAF étaient disponibles pour 60 lincRNA uniques à Sapiens (tableau supplémentaire S6) et 53 présents chez les Néandertaliens (tableau supplémentaire S7), ce qui nous a permis de les comparer (tableau supplémentaire S12). Parmi les lincRNAs uniques à Sapiens (Supplementary Fig. S4C), ceux qui ont un DAF > 0.1 sont plus fréquents que ceux qui ont un DAF plus faible à la fois pour leurs promoteurs (F (1,110) = 30.23, p < 0.0001) et leurs exons (F (1,110) = 9.78, p < 0.0022). De même, les promoteurs (F (1,92) = 45,35, p < 0,0001) et les exons (F (1,94) = 11,75, p < 0,0019) sont principalement soumis à une sélection positive dans les lincRNA qui sont également présents chez les Néandertaliens. Cependant, les promoteurs de lincRNA chez Sapiens, mais pas chez Neandertal, ont DAF > 0,1 un peu plus souvent que leurs exons (F (1,102) = 4,54, p < 0,03), ce qui suggère que la sélection positive agit sur les fonctions de régulation chez Sapiens. De manière significative, ~70% des 49 lincRNAs uniques à Sapiens et soumis à une sélection positive (DAF > 0,1) se trouvaient dans le réseau génotypique de la conscience de soi.
Expression d’ensembles de gènes propres aux modernes dans des régions spécifiques du cerveau
94 % des 267 gènes uniques à l’homme étaient des gènes codant pour des protéines, et 64 % étaient associés au réseau génotypique de la conscience de soi (tableau supplémentaire S5). Leurs fonctions spécifiques sont largement incertaines, si ce n’est qu’en général, il est suggéré qu’ils coordonnent des processus complexes d’adaptation, de plasticité et de santé en régulant la co-expression de groupes d’autres gènes. Par conséquent, nous avons évalué la co-expression dans différentes régions du cerveau des ensembles de gènes qui correspondaient au même ensemble de SNP liés à la personnalité et qui contenaient au moins un gène trouvé uniquement chez Sapiens (méthodes supplémentaires et tableau S13). Nous avons calculé le niveau d’expression moyen de l’ARNm dans des régions cérébrales spécifiques des groupes multigéniques liés au caractère ou au tempérament.
94 % des 267 gènes uniques à l’homme étaient des gènes codant pour des protéines, et 64 % étaient associés au réseau génotypique de la conscience de soi (tableau supplémentaire S5). Leurs fonctions spécifiques sont largement incertaines, si ce n’est qu’en général, il est suggéré qu’ils coordonnent des processus complexes d’adaptation, de plasticité et de santé en régulant la co-expression de groupes d’autres gènes. Par conséquent, nous avons évalué la co-expression dans différentes régions du cerveau des ensembles de gènes qui correspondaient au même ensemble de SNP liés à la personnalité et qui contenaient au moins un gène trouvé uniquement chez Sapiens (méthodes supplémentaires et tableau S13). Nous avons calculé le niveau d’expression moyen de l’ARNm dans des régions cérébrales spécifiques des groupes multigéniques liés au caractère ou au tempérament.
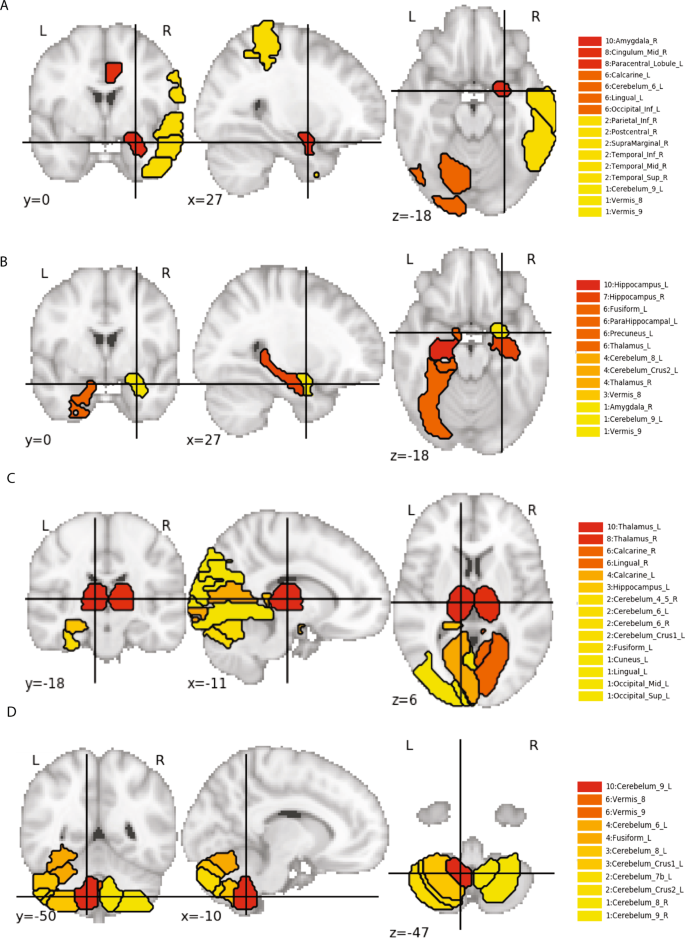
En outre, nous avons constaté que les gènes liés au tempérament étaient significativement surexprimés dans l’hippocampe, le septum, l’amygdale, le cortex cingulaire, le gyrus parahippocampique, le gyrus fusiforme, le thalamus, le cervelet et le tronc cérébral (Fig. 3C, D, Tableau supplémentaire S14), comme prévu en raison du rôle de ces régions dans la reconnaissance, l’évaluation et l’autorégulation de l’expression émotionnelle.
Lorsqu’ils sont associés à des gènes trouvés uniquement chez Sapiens, les gènes des réseaux de la conscience de soi et du contrôle de soi sont significativement surexprimés dans les régions du cerveau qui comprennent les voies de la conscience de soi et de l’autorégulation des émotions et des objectifs. En d’autres termes, la régulation de la co-expression des gènes par des gènes trouvés uniquement chez Sapiens a fourni un mécanisme permettant d’intégrer la conscience de soi et le contrôle de soi, de sorte que les émotions, les objectifs et les valeurs humaines puissent être autogérés de manière cohérente, raisonnable et avantageuse.
Discussion
Il s’agit de la première étude visant à identifier les différences génotypiques entre les chimpanzés, les Néandertaliens et les humains modernes qui peuvent expliquer l’émergence rapide de la créativité humaine et d’autres composantes de la modernité comportementale, y compris ses caractéristiques physiques, émotionnelles, cognitives, sociales et spirituelles. Lors de travaux préparatoires, nous avons identifié trois réseaux génotypiques naturels pour la réactivité émotionnelle, l’autocontrôle intentionnel et la conscience de soi. Les 972 gènes de ces réseaux expliquent la quasi-totalité de la variation héréditaire de la personnalité humaine, y compris les caractéristiques de la modernité comportementale (à savoir, la créativité, le comportement prosocial et la longévité en bonne santé). Nous avons découvert que 267 de ces gènes sont absents des génomes des chimpanzés et des Néandertaliens, et nous avons reproduit cette découverte dans trois génomes néandertaliens à forte couverture.
Nous avons également constaté que les Néandertaliens avaient presque les mêmes proportions de gènes de réactivité émotionnelle que les chimpanzés. Si l’on exclut 54 gènes présents uniquement chez Sapiens, 72 % des 195 gènes de la réactivité émotionnelle étaient communs aux trois espèces. En revanche, les Néandertaliens se situaient à un niveau intermédiaire par rapport aux chimpanzés et aux Sapiens en ce qui concerne les proportions de gènes de contrôle de soi et de conscience de soi. Si l’on fait abstraction des gènes de la personnalité présents chez les chimpanzés, les Néandertaliens possédaient 33 % des gènes de la conscience de soi et 37 % des gènes du contrôle de soi présents chez les Sapiens. Néanmoins, en tenant compte de l’organisation modulaire de ces gènes en grappes avec d’autres gènes, nous avons estimé que le bien-être relatif des humains de type néandertalien représentait 61 à 70 % de celui des Sapiens prototypiques qui portaient des gènes présents uniquement chez les humains modernes. Les Sapiens prototypiques ont une prédisposition génotypique beaucoup plus forte aux caractéristiques de la modernité comportementale que les humains de type Néandertal, en particulier des ensembles de gènes du réseau de la conscience de soi associés à la créativité, au comportement prosocial et à la longévité (F (3,252), p < 00001, taille de l’effet de Cohen f = 0,34).
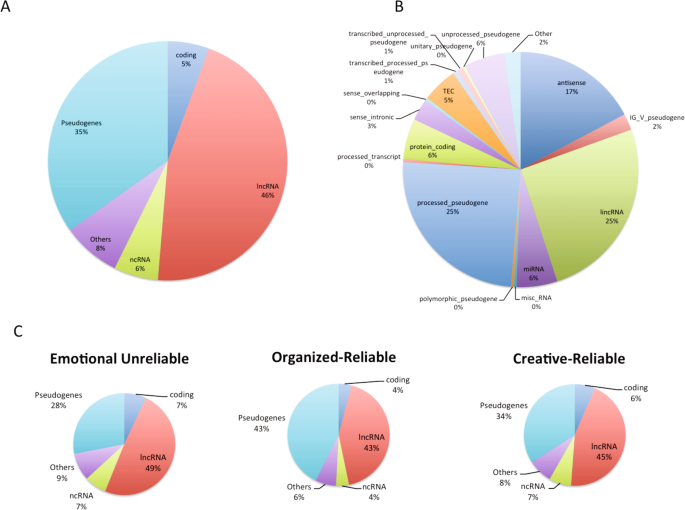
En outre, nous avons obtenu des preuves que les gènes trouvés uniquement chez Sapiens étaient probablement régulateurs et avantageux. Plus précisément, 94 % des 267 gènes trouvés uniquement chez Sapiens n’étaient pas codants pour les protéines, notamment de nombreux lncRNA (46 %), pseudogènes (35 %) et ncRNA (6 %). 64 % des gènes trouvés uniquement chez Sapiens faisaient partie du réseau de la conscience de soi, en particulier les lncRNA que nous avons trouvés soumis à une sélection positive.
Enfin, nous avons testé l’importance des gènes uniques à Sapiens pour le bien-être humain et la modernité comportementale en identifiant les régions du cerveau dans lesquelles ils étaient surexprimés. Nous avons confirmé que des groupes de gènes naturels associés à un ou plusieurs gènes trouvés uniquement chez Sapiens étaient surexprimés dans les régions du cerveau essentielles à la conscience de soi, qui est fortement associée au bien-être humain, y compris les caractéristiques identifiées par les anthropologues comme distinguant Sapiens des autres hominidés qu’ils ont remplacés vers 40 kya.
En gardant ces résultats clés à l’esprit, nous discuterons des données anthropologiques et génétiques disponibles pour tester nos hypothèses liées à l’émergence successive de réseaux presque disjoints pour la régulation de la réactivité émotionnelle, le contrôle intentionnel de soi et la conscience créative de soi dans la lignée hominoïde des humains modernes. D’après nos études préparatoires de l’architecture phénotypique-génotypique de la personnalité humaine, nous reconnaissons que ces trois réseaux fonctionnent de manière coopérative afin qu’une personne puisse apprendre à intégrer ses habitudes, ses objectifs et ses valeurs en s’adaptant aux changements de son milieu interne et externe. Les informations disponibles sur les changements coïncidents dans le fonctionnement du cerveau et du comportement dans la phylogénie de Sapiens aident à guider notre interprétation de nos résultats basés sur la comparaison des génomes des chimpanzés, des Néandertaliens et de Sapiens.
Émergence du réseau de régulation des émotions sociales
Les ancêtres mammifères des primates anthropoïdes étaient pour la plupart de petite taille, nocturnes et solitaires ; mais lorsque les températures se sont refroidies et que les forêts tropicales ont reculé à la fin de l’Éocène, il y a environ 40 millions d’années (mya), il y a probablement eu un avantage sélectif dans la coopération sociale chez les primates supérieurs comme protection contre les prédateurs lors de la recherche de nourriture pendant la journée [12, 14]. Un apprentissage social similaire à celui de l’homme s’est ensuite développé chez les singes et les grands singes, aboutissant à l’attachement social [98, 99], à la régulation de la réactivité émotionnelle en fonction du contexte social et à la réduction de la détresse émotionnelle par la réconciliation [100], comme chez les chimpanzés d’aujourd’hui qui, après un combat, pratiquent souvent le bouche-à-bouche et les étreintes ventrales. L’apprentissage social permet également la transmission proto-culturelle des traditions de toilettage, de cour, de recherche de nourriture et de préparation des aliments [101,102,103]. Les gestes émotionnels et les appels vocaux facilitent les relations sociales au sein des triades et des groupes plus importants de primates supérieurs, de sorte qu’un tiers, tel qu’un chef de groupe de haut rang, peut intervenir pour résoudre les conflits [100, 104].
D’autre part, si les chimpanzés font preuve d’une réactivité émotionnelle et de capacités d’apprentissage similaires à celles d’un enfant humain moderne de 2 ou 3 ans, ils ne présentent pas les capacités de régulation des enfants humains modernes plus âgés [105]. Les chimpanzés utilisent des outils pour résoudre des tâches simples, comme casser des noix ou attraper des termites, mais ils ne s’apprennent pas mutuellement à fabriquer et à utiliser ces outils [1]. On peut leur apprendre à utiliser des signes et à former des phrases de deux à quatre mots à un rythme compatible avec le conditionnement comportemental, mais, contrairement aux enfants humains modernes, ils n’acquièrent pas spontanément le langage symbolique [45, 106, 107]. La mémoire consciente de soi des enfants humains modernes commence à mûrir vers l’âge de 4 ans, et par la suite ils montrent une plus grande capacité que les chimpanzés à retarder la gratification, à raisonner sur les croyances et à résoudre des problèmes concernant les souvenirs internes [57, 105, 106, 107, 108].
Lorsque l’on compare le cerveau des primates supérieurs à celui de parents plus éloignés de l’homme [12, 45], le cortex préfrontal est typiquement hypertrophié, se projetant directement sur l’hypothalamus, le striatum, le thalamus, le septum et l’amygdale basale. Les informations affectives sont également relayées vers le cortex insulaire moyen, qui permet de réguler la sensualité. Le système des neurones miroirs émerge, permettant la compréhension de l’action et l’imitation des comportements observés, précurseur nécessaire du langage. Chez les grands singes, on observe également une différenciation du cortex insulaire antérieur, permettant une conscience émotionnelle accrue qui favorise la communication des émotions sociales. Sur la base de ces constatations de changements coïncidents dans le cerveau et le comportement, nous avons émis l’hypothèse que le génome des chimpanzés est susceptible de posséder le réseau génétique pour la régulation de la réactivité émotionnelle, mais pas ceux pour le contrôle intentionnel de soi ou la conscience créative de soi [12, 45]. Nos résultats actuels confirment fortement cette hypothèse : le réseau de la réactivité émotionnelle est bien développé chez les trois espèces d’hominoïdes que nous avons évaluées. Si l’on met de côté les 54 gènes trouvés uniquement chez Sapiens, 72 % des 195 gènes du réseau de réactivité émotionnelle étaient partagés par les trois espèces (tableau 1).
Emergence du réseau de régulation de l’autocontrôle intentionnel
Les premiers hominines se sont rapidement distingués des grands singes par une plus grande facilité à adopter des comportements de recherche de but précis, comme la fabrication d’outils et la chasse coordonnée pour la nourriture [12, 45]. Selon les indications actuelles, l’utilisation et la fabrication d’outils en pierre ont été introduites par des ” australopithèques ” aux proportions archaïques (par exemple, [109]) à une époque où les habitats ouverts se répandaient à mesure que les forêts tropicales se réduisaient. Par la suite, la possession de proportions de membres plus ou moins modernes par les premiers membres correctement diagnostiqués du genre Homo indique que les hominines se sont finalement engagés dans ces habitats ouverts il y a un peu moins de 2 millions d’années. Cette transition cruciale est mal documentée en termes de comportement, mais elle a certainement représenté un changement environnemental et économique extrême qui a dû avoir de profondes séquelles cognitives et sociales.
Une fois engagés dans des habitats ouverts, la taille du cerveau des hominines a commencé à augmenter rapidement. L’Homo ergaster (littéralement, l’homme qui travaille) était raisonnablement grand et svelte, comme les humains modernes, et a introduit l’industrie de l’outillage acheulien des haches à main bifaciales symétriques avant 1,6 mya. Ces outils étaient intentionnellement écaillés pour se conformer à un modèle que leurs fabricants avaient en tête. Les hominines ultérieures ont poursuivi cette tradition de fabrication d’outils sans innovation radicale jusqu’à environ 400 kya [9, 10]. Cet enregistrement archéologique de la stase technologique pendant plus d’un million d’années prouve que les premiers humains avaient la capacité de se contrôler intentionnellement, mais que les humains vivant avant 400 kya, y compris l’ancêtre commun des Néandertaliens et des Sapiens, ne manifestaient pas la créativité associée au réseau génotypique de la conscience de soi des Sapiens [12].
L’Homo neanderthalensis, une espèce qui a évolué à partir d’un précurseur européen endémique il y a environ 200 000 ans, est un produit final très évolué de l’engagement humain à vivre dans des habitats ouverts. Les Néandertaliens étaient manifestement des créatures déterminées et pleines de ressources qui vivaient généralement en petits groupes de 12 à 25 individus qui s’alimentaient dans de vastes paysages [110]. Il s’agissait manifestement d’êtres sophistiqués, très opportunistes quant aux ressources qu’ils exploitaient : ils chassaient des proies d’une taille effrayante lorsque les circonstances l’exigeaient (ce qui pourrait expliquer l’incidence élevée de fractures osseuses signalée [110]) ; au moins occasionnellement, ils construisaient des abris et contrôlaient le feu dans des foyers [111,112,113]. Dans la grotte de Shanidar, dans le nord de l’Irak, on a trouvé la preuve qu’un Néandertalien a survécu jusqu’à un âge avancé malgré le fait qu’il était gravement handicapé par un bras inutile, ce qui suggère une coopération sociale et une empathie pour les autres au sein de leurs petits groupes [113]. D’autre part, si les Néandertaliens enterraient leurs morts, ils le faisaient généralement sans les objets funéraires caractéristiques des sépultures ultérieures des Cro-Magnons [113, 114]. Les Néandertaliens ont produit des artefacts qui ont été interprétés comme de l’art symbolique, mais ces expressions peu fréquentes étaient simples et bidimensionnelles [73, 74, 75], probablement comparables aux images produites par les enfants humains modernes avant l’âge de 7 ans [115]. Leur faible diversité génétique suggère qu’ils vivaient en petits isolats avec des accouplements limités entre les groupes [110, 116], bien qu’il existe des preuves d’exogamie féminine [115].
Au cours de la période qui a suivi, il y a 40 000 ans, les Néandertaliens ont été rapidement remplacés en Europe, bien qu’il y ait eu quelques échanges génétiques mineurs [117], par des Homo sapiens envahisseurs dont la vie présentait une sophistication culturelle et technologique sans précédent. Alors qu’ils étaient encore des chasseurs-cueilleurs itinérants, ces nouveaux humains, qui se distinguaient par leur anatomie et leur comportement, ont peuplé le paysage en plus grandes densités et ont apporté avec eux la tradition symbolique de l’art rupestre narratif, qui utilise des indices de profondeur picturale dans des compositions intégrées d’une grande complexité et d’une grande beauté [118]. Cette pratique innovante consistant à créer des images à partir de l’imagination – “l’œil de l’esprit” – est l’indicateur le plus puissant que nous ayons de l’éveil de la sensibilité moderne, avec sa profusion de signes abstraits mais clairement chargés de sens, en plus des images animales sophistiquées célèbres dans des localités comme Chauvet et Lascaux [73].
Les cerveaux des humains éteints ne sont disponibles que sous forme d’endocastes fossiles, ce qui limite les observations qui peuvent être faites. Par rapport aux chimpanzés, les données fossiles documentent l’émergence de l’asymétrie hémisphérique ainsi que de la bipédie chez les australopithèques et les non-Sapiens. Apparus tardivement dans l’histoire des hominidés, les Néandertaliens avaient un gros cerveau d’un volume moyen d’environ 1500 ml, plus ou moins identique à celui des Homo sapiens contemporains du Pléistocène (bien que le cerveau des humains modernes soit plus petit de près de 13% [117]). Cependant, ces cerveaux semblent avoir été organisés différemment des cerveaux modernes : Les Néandertaliens avaient des zones visuelles relativement plus grandes, tandis que les Sapiens ont des lobes pariétaux élargis [69] et des régions préfrontales plus élevées. Sur la base de ces constatations de différences coïncidentes dans le cerveau et le comportement, nous avons émis l’hypothèse que le génome des Néandertaliens comporterait probablement le réseau génétique de régulation de la réactivité émotionnelle et certains des gènes du réseau d’autocontrôle intentionnel, mais pas celui de la conscience de soi [12, 45].
Nos résultats actuels confirment que le réseau génotypique pour le contrôle intentionnel de soi est bien développé chez les Néandertaliens mais pas chez les chimpanzés. Ils suggèrent également que les Néandertaliens ont acquis des gènes pour le contrôle de soi et la conscience de soi en nombre intermédiaire entre l’homme moderne et le chimpanzé. Si l’on exclut les gènes déjà présents chez les chimpanzés, les Néandertaliens possédaient 33 % des 254 gènes de la conscience de soi et 37 % des 186 gènes du contrôle de soi présents chez Sapiens. En tenant compte de l’organisation modulaire des groupes de gènes dans les réseaux d’apprentissage humains, nous avons estimé que le niveau relatif de prédisposition génotypique au bien-être et à la modernité des humains de type néandertalien représentait 61 à 70 % de celui des Sapiens prototypiques. Par rapport au Sapiens prototypique, la prédisposition génotypique à la modernité des humains de type Néandertal est la plus faible pour la conscience de soi (taille de l’effet de Cohen f = 0,34). Ces résultats suggèrent que l’événement crucial qui a déclenché l’émergence de la modernité comportementale a été l’évolution avancée du réseau génotypique de la conscience de soi chez Sapiens, mais nous devons envisager d’autres explications pour ces résultats.
Bien sûr, une autre explication possible est que tous les gènes présents chez les Néandertaliens n’ont peut-être pas été documentés dans les informations génomiques dont nous disposons actuellement, même si nous avons reproduit nos résultats en utilisant le projet de génome de 2010 séparément dans chacun des trois génomes néandertaliens à couverture élevée qui sont disponibles : Vindija 33.19 de l’aire centrale des Néandertaliens en Croatie, ainsi que les génomes d’un Néandertalien des montagnes de l’Altaï et d’un autre de la grotte de Chagyrskaya en Russie [110, 116, 117, 119]. Ces résultats répétés ont apporté un soutien solide à nos analyses comparatives, mais nous devions encore savoir si les gènes que nous avions trouvés fournissaient un mécanisme pouvant expliquer l’émergence de la créativité.
Émergence du réseau pour la conscience créative de soi
Quel mécanisme a favorisé l’émergence du réseau génétique de la conscience créative de soi chez les êtres humains modernes sur le plan comportemental ? Le cerveau de Sapiens est unique car il possède un système de conscience de soi qui relie les régions à myélinisation tardive des cortex frontal, pariétal et temporal [57, 120]. Ces régions les plus récemment évoluées du cerveau sont les dernières zones d’association dans lesquelles les informations sont intégrées et évaluées, et sont reliées à un réseau unifié pour la mémoire épisodique par des projections du cortex visuel [12, 45]. L’apprentissage et la mémoire autobiographiques permettent de prendre conscience du soi en tant qu’identité continue dans l’espace et le temps. Sur le plan psychologique, le réseau créatif est ainsi nommé parce qu’on le trouve chez les personnes imaginatives, inventives, prosociales et spirituelles [42, 47, 48, 55, 80, 121]. Cette pensée autotranscendante implique la capacité de se percevoir comme un aspect local d’un ensemble spatio-temporel plus vaste, ce qui permet une pensée libre et créative (c’est-à-dire “out of the box” de la déduction logique et de la tradition culturelle) et théoriquement inductive (c’est-à-dire une extrapolation au-delà des exemples antérieurs basée sur la perspicacité et l’imagination créative), telle qu’elle s’exprime dans l’art, la science, la spiritualité et le langage syntaxique narratif [12, 45]. Sur la base des résultats de l’association unique de changements cérébraux coïncidents avec les fonctions cognitives de la conscience de soi et de la créativité, nous avons émis l’hypothèse que seul Sapiens était susceptible de posséder le réseau génotypique de la conscience de soi.
Cependant, cette hypothèse n’a été que partiellement confirmée. Nous avons constaté que les Néandertaliens ne possédaient que 33 % des gènes de la conscience de soi présents chez les Sapiens ; mais ces gènes, lorsqu’ils sont organisés en grappes avec d’autres gènes humains, sont suffisants pour que les humains de type Néandertalien fonctionnent à 61-70 % du niveau de bien-être des Sapiens prototypiques. Cela ne nous permet toujours pas de savoir si les Néandertaliens avaient franchi le seuil génotypique nécessaire pour avoir le potentiel d’exprimer tout ou partie des caractéristiques de la modernité comportementale, même si cette capacité n’a pas été suffisamment documentée dans les archives archéologiques.
En outre, les caractéristiques d’altruisme et de longévité en bonne santé peuvent avoir fourni les conditions nécessaires à la sélection de la parenté pour la créativité chez Sapiens en tant que réponse adaptative à la pression écologique intense due aux fluctuations climatiques et à la variabilité imprévisible de la disponibilité des ressources en Afrique de l’Est, mais pas chez les Néandertaliens qui n’étaient pas soumis aux mêmes pressions en Europe. L’importance des environnements prosociaux pour la réalisation de la créativité est encore évidente dans les différences comportementales observées aujourd’hui chez les humains modernes : même les Sapiens dotés du réseau génotypique de la conscience de soi sont encore vulnérables au mal-être physique, émotionnel, cognitif et social dans des conditions sociales hostiles ou inéquitables [19], comme le montre la figure S5D. Par conséquent, les comportements altruistes et créatifs sont des caractéristiques fréquentes, mais incohérentes, de Sapiens [121, 126].
Compte tenu de toutes les preuves disponibles, nous savons que les Néandertaliens étaient intermédiaires entre les chimpanzés et les Sapiens dans le développement du réseau génotypique de la conscience de soi. Nous savons également que les Sapiens possèdent un ensemble distinctif de gènes qui se trouvent principalement dans le réseau de la conscience de soi, qui sont soumis à une sélection positive et qui ne sont pas présents chez les Néandertaliens. Nos résultats génotypiques documentent des mécanismes moléculaires qui peuvent fournir une explication probable aux archives archéologiques qui n’ont trouvé que des preuves rudimentaires de créativité et d’autres signes de modernité comportementale chez les Néandertaliens. Nous devons donc examiner en détail ces mécanismes potentiellement cruciaux.
Hypothèses sur la sélection pour la créativité
La créativité nouvellement apparue peut avoir procuré des avantages sélectifs aux humains modernes sur le plan comportemental, au-delà de ses avantages purement cognitifs. Sur le plan physiologique, elle est associée à une amélioration des fonctions de la mémoire, de la santé et du bien-être (figures supplémentaires 5 et 6), y compris une prédisposition à la longévité et à la résistance au stress, aux blessures et aux maladies chroniques, notamment les maladies cardiovasculaires et neurodégénératives [19, 64, 65]. Le fait de vivre plus longtemps et en meilleure santé a peut-être permis à l’Homo sapiens moderne sur le plan comportemental de se disperser rapidement et largement dans le monde, et a peut-être aussi aidé les individus à soutenir leurs enfants, petits-enfants et autres dans des communautés sociales interconnectées, ce qui a peut-être entraîné une sélection positive pour des traits tels que la créativité, l’innovation, la prosocialité et la sagesse [127,128,129,130,131]. Nous avons émis l’hypothèse que le réseau génétique de la créativité a fait l’objet d’une sélection positive, car nous avions précédemment constaté que la longévité et le bien-être sont favorisés par l’intégration du fonctionnement créatif, de la plasticité et de vertus telles que la modération, l’altruisme et la sagesse [19]. Cette hypothèse est également soutenue par notre constatation que 70 % des lncRNA avantageux propres à Sapiens se trouvaient dans le réseau de la conscience de soi, qui est fortement associé à la créativité, à la prosocialité et à une longévité saine [19, 55, 80]. Il est donc crucial de constater que la plupart des gènes régulateurs clés de la conscience créative de soi ne sont présents que chez Sapiens, et non chez les Néandertaliens : sur les 130 lncRNA du réseau de la conscience de soi, aucun n’était présent chez les chimpanzés, 42 % étaient partagés par Sapiens et les Néandertaliens, et 58 % n’étaient présents que chez les humains modernes (tableau 1, figure 2, tableau supplémentaire S3).
Rôle des LncRNAS dans les changements rapides de l’évolution
Quel mécanisme peut expliquer la rapidité de l’évolution de la créativité, de la longévité en bonne santé et de la forme physique chez Sapiens [1,2,3,4, 132] ? Les changements dans les taux de mutation ne fournissent pas d’explication car ils sont restés stables lors de la transition de l’homme archaïque à l’homme moderne [119, 133]. Nous avons envisagé des mécanismes par lesquels de nouveaux gènes apparaissent sans dépendre du taux de mutation des gènes ancestraux [134]. Nous avons observé que 67% des gènes associés à l’autorégulation et à la créativité humaine étaient des gènes régulateurs [64], incluant une prédominance significative de gènes lncRNA et de pseudogènes par rapport aux gènes liés au conditionnement comportemental du tempérament [65]. Nous savons que les différences de complexité des fonctions entre les espèces dépendent généralement de différences dans la régulation de l’expression génétique d’un noyau hautement conservé de gènes codant pour des protéines, comme cela a été démontré pour les différences entre les chimpanzés et les humains [135,136,137].
Plus précisément, nous savons que les lncRNA sont souvent des régulateurs importants de l’expression des gènes [138] et qu’ils sont souvent acquis par transfert horizontal de gènes (THG) [88]. Le transfert horizontal de gènes (c’est-à-dire l’acquisition de gènes d’un organisme autre qu’un ancêtre direct) permet aux génomes de se développer rapidement, d’assembler de nouvelles voies et d’exprimer de nouvelles fonctions [139]. Le HGT est le principal mécanisme d’acquisition de nouveaux gènes chez les procaryotes et les eucaryotes unicellulaires, et il est également très répandu chez les primates, y compris les humains. De nombreux nouveaux gènes ont été acquis dans l’ensemble du génome humain moderne, en particulier des gènes codant pour des protéines et des lncRNA (par exemple, lincRNA et antisens) [88]. Nous avons donc testé l’hypothèse selon laquelle les êtres humains modernes ont acquis les gènes qui ont permis l’évolution rapide de la créativité et de la longévité en bonne santé par HGT. Nous avons constaté que les gènes de la personnalité humaine sont enrichis dans les régions HGT, mais l’enrichissement a été observé pour les gènes du réseau de réactivité émotionnelle ainsi que pour les autres. En outre, seuls 2 des 39 gènes que nous avons trouvés dans les régions HGT étaient uniques aux humains modernes. Par conséquent, nous avons conclu que le HGT peut avoir contribué au développement de la personnalité chez les hominoïdes en général, mais qu’il n’a pas joué un rôle majeur dans le développement de la personnalité créative ou de la conscience de soi.
En revanche, notre constat que 70% des lincRNAs uniques aux humains et sous sélection positive se trouvaient exclusivement dans le réseau de la conscience de soi fournit des preuves de leur implication dans l’évolution de la conscience de soi et des divers aspects du bien-être humain et de la modernité comportementale. De même, 35% des gènes uniques à Sapiens étaient des pseudogènes, qui sont également souvent soumis à une sélection positive chez les primates [140, 141] et impliqués dans la régulation de la cognition humaine [142]. Les pseudogènes étaient plus fréquents dans les gènes associés à la personnalité chez Sapiens (8% sur 972) que chez Neandertal (2% sur 652). Cependant, chez Sapiens, les pseudogènes étaient plus fréquents dans le réseau de la maîtrise de soi (43 %) que dans celui de la conscience de soi (28 %). Par conséquent, les lncRNA semblent avoir joué un rôle plus direct dans l’émergence de la créativité chez Sapiens, bien que les pseudogènes contribuent également de manière substantielle aux différences entre les deux espèces humaines qui ont émergé dans des conditions écologiques distinctes.
Contrairement aux différences que nous avons observées dans les biotypes entre les espèces, nous avons constaté que les biotypes des gènes sont similaires pour chacun des trois réseaux au sein de chaque espèce (Fig. 1). En somme, tant les différences de biotypes entre les espèces que la similarité des biotypes entre les réseaux adaptatifs au sein des espèces soutiennent notre hypothèse selon laquelle les réseaux génotypiques presque disjoints sont susceptibles d’avoir émergé par étapes progressives. L’émergence initiale de la fixation intentionnelle d’objectifs chez les premiers hominines et, plus tard, l’émergence de l’imagination créative de Sapiens ont permis aux humains modernes de s’adapter aux défis sociaux et environnementaux grâce à des fonctions cérébrales qui sont associées à des processus moléculaires distinctifs et à de nombreux gènes régulateurs que l’on retrouve chez les humains modernes, mais pas chez les chimpanzés ou les Néandertaliens.
Points forts et limites
La principale innovation et la force de notre étude de l’évolution de la créativité humaine est d’avoir commencé par caractériser l’architecture génotypique-phénotypique complexe de la personnalité humaine qui sous-tend la capacité humaine de conscience de soi, de symbolisme et de créativité. Nous avons identifié et reproduit les réseaux génotypiques qui sous-tendent les trois principaux systèmes d’apprentissage chez Sapiens (conditionnement comportemental, intentionnalité et conscience de soi). Cela nous a permis de concentrer les analyses génomiques comparatives sur 972 gènes qui expliquent la personnalité humaine moderne et les capacités d’apprentissage.
Le fait qu’il existe moins d’informations sur le génome de l’homme de Neandertal que sur celui de l’Homo sapiens et du chimpanzé modernes a constitué un défi majeur. Le génome annoté du Neanderthal Genome Project de 2010 est basé sur des données à faible couverture, dont la quasi-totalité provient de la grotte de Vindija en Croatie, située dans la zone centrale de l’aire de répartition des Néandertaliens pendant la majeure partie de leur existence. Heureusement, nous avons pu reproduire nos premiers résultats avec le génome complet à haute couverture (~50×) de l’homme de Neandertal de l’Altaï, qui a confirmé les mêmes 267 gènes de Sapiens qui étaient absents chez les Néandertaliens de Vindija. Nos résultats ont également été confirmés par des analyses séparées de deux autres génomes à haute couverture (~30×) provenant de grottes situées en Croatie et en Russie, de sorte que nos résultats sont solides.
Comme nous nous sommes concentrés uniquement sur les 972 gènes qui expliquent la personnalité chez Sapiens, nous ne pouvons pas exclure la possibilité que les Néandertaliens possédaient des gènes qui n’étaient pas présents chez Sapiens et qui ont influencé leur personnalité et leurs capacités d’apprentissage. Ces gènes pourraient avoir été hérités de l’ancêtre commun des Néandertaliens et des Sapiens ou acquis par les Néandertaliens ultérieurement. Tous ces gènes uniques de Neandertal pourraient avoir eu des fonctions homologues ou distinctes de celles présentes chez les humains modernes. Cependant, nous avons identifié les gènes présents chez Sapiens, mais pas chez Neandertal, qui expliquent l’émergence des capacités avantageuses de Sapiens, notamment la conscience créative de soi, le comportement prosocial et la longévité. Les données comportementales disponibles indiquent également que ces mêmes capacités étaient absentes chez les Néandertaliens et d’autres hominidés éteints, et des analyses génotypiques-phénotypiques plus détaillées comparables à ce que nous avons fait chez les humains modernes sont impossibles. Par conséquent, il sera probablement beaucoup plus utile de chercher à comprendre plus en détail les fonctions des gènes propres à Sapiens que celles des gènes propres aux hominidés disparus.
Un autre défi majeur était le peu d’informations connues sur les fonctions des gènes d’ARN non codants qui constituaient la plupart des gènes présents uniquement chez Sapiens. Heureusement, il a été démontré que les gènes lncRNA régulent l’expression d’ensembles d’autres gènes. Nous avons donc pu identifier les régions cérébrales spécifiques dans lesquelles s’expriment les génotypes multilocus qui correspondent aux ensembles de SNP liés à la conscience de soi chez Sapiens. Nos résultats concernant l’expression des gènes dans le cerveau du réseau de la conscience de soi ont confirmé les résultats de l’imagerie cérébrale fonctionnelle concernant les régions du cerveau impliquées dans diverses fonctions de la conscience de soi, notamment la mémoire autobiographique, la prospection, la théorie de l’esprit et le mode par défaut [59]. Nos résultats les ont étendus en révélant des structures sous-corticales supplémentaires qui sont impliquées dans les boucles de rétroaction corticales importantes pour le traitement et l’intégration automatiques des informations dans la conscience de soi. La reproductibilité de nos résultats génétiques et leur association significative avec des circuits cérébraux spécifiques pour des fonctions humaines complexes constituent des preuves solides de la validité des méthodes basées sur les données que nous avons développées et appliquées pour caractériser les systèmes adaptatifs complexes [64].
Vue d’ensemble
Nos résultats ont de vastes implications pour comprendre ce qui a permis à Sapiens de déplacer les Néandertaliens et d’autres espèces d’Homo dans le passé géologiquement récent, ainsi que de remodeler littéralement le monde au cours de l’Anthropocène. Le fait de vivre plus longtemps et en meilleure santé peut avoir favorisé et valorisé les périodes prolongées d’apprentissage chez les jeunes et les adolescents qui permettent l’accumulation de connaissances qui est une caractéristique si remarquable des humains modernes sur le plan comportemental, et qui est un facteur si important dans la réussite économique et les structures et relations sociales complexes d’Homo sapiens [145]. Il est également possible qu’elle ait encouragé la coopération entre les individus afin de promouvoir la réussite de leurs enfants, petits-enfants et autres membres de leur communauté élargie [128, 131], permettant ainsi l’innovation technologique, la flexibilité comportementale et la disposition à l’exploration nécessaires pour permettre à l’Homo sapiens de se répandre dans le monde avec plus de succès que d’autres lignées humaines [1, 2, 3]. D’autres travaux sont nécessaires pour comprendre les fonctions spécifiques des lncRNA associés à la conscience de soi qui sous-tendent la capacité des humains modernes à jouir d’une longévité saine, de la prosocialité et de la créativité. Une compréhension plus complète est grandement nécessaire en raison de l’échec fréquent de ces capacités bénéfiques des humains modernes à s’accomplir au cours de l’Anthropocène [52].
Disponibilité des données
L’étude Young Finns a accordé l’accès aux données au CRC et à IZ par un accord de transfert de matériel.
Références
- 1.Paabo S. The human condition-a molecular approach. Cell. 2014;157:216–26.PubMed Article CAS PubMed Central Google Scholar
- 2.Tattersall I. Out of Africa: modern human origins special feature: human origins: out of Africa. Proc Natl Acad Sci USA. 2009;106:16018–21.CAS PubMed Article PubMed Central Google Scholar
- 3.Mellars P. Going east: new genetic and archaeological perspectives on the modern human colonization of Eurasia. Science. 2006;313:796–800.CAS PubMed Article PubMed Central Google Scholar
- 4.Mellars P, Gori KC, Carr M, Soares PA, Richards MB. Genetic and archaeological perspectives on the initial modern human colonization of southern Asia. Proc Natl Acad Sci USA. 2013;110:10699–704.CAS PubMed Article PubMed Central Google Scholar
- 5.Klein RG. The human career: human biological and cultural origins. 3rd edn. University of Chicago Press: Chicago, 2009.
- 6.Tattersall I. An evolutionary framework for the acquisition of symbolic cognition by Homo sapiens. Comp Cognition Behav Rev. 2008;3:99–114.Google Scholar
- 7.Hauser MD, Yang C, Berwick RC, Tattersall I, Ryan MJ, Watumull J, et al. The mystery of language evolution. Front Psychol. 2014;5:401.PubMed PubMed Central Google Scholar
- 8.Elias S. (ed). Origins of human innovation and creativity. Elsevier: Amsterdam, 2012.
- 9.Tattersall I. Masters of the planet: the search for our human origins. St. Marti’s Press, 2012.
- 10.Lopez S, van Dorp L, Hellenthal G. Human dispersal out of Africa: a lasting debate. Evol Bioinform Online. 2015;11:57–68.PubMed PubMed Central Google Scholar
- 11.Hill K, Barton M, Hurtado AM. The emergence of human uniqueness: characters underlying behavioral modernity. Evolut Anthropol. 2009;18:187–200.Article Google Scholar
- 12.Cloninger CR. The evolution of human brain functions: the functional structure of human consciousness. Aust NZ J Psychiatry. 2009;43:994–1006.Article Google Scholar
- 13.Suddendorf T, Addis DR, Corballis MC. Mental time travel and the shaping of the human mind. Philos Trans R Soc Lond B Biol Sci. 2009;364:1317–24.PubMed PubMed Central Article Google Scholar
- 14.Sussman RW, Cloninger CR. (eds). Origins of cooperation and altruism. Springer: New York, 2011.
- 15.Brooks AS, Yellen JE, Potts R, Behrensmeyer AK, Deino AL, Leslie DE, et al. Long-distance stone transport and pigment use in the earliest Middle Stone Age. Science. 2018;360:90–94.CAS PubMed Article PubMed Central Google Scholar
- 16.Staubwasser M, Dragusin V, Onac BP, Assonov S, Ersek V, Hoffmann DL, et al. Impact of climate change on the transition of Neanderthals to modern humans in Europe. Proc Natl Acad Sci USA. 2018;115:9116–21.CAS PubMed Article PubMed Central Google Scholar
- 17.Anton SC, Potts R, Aiello LC. Evolution of early Homo: an integrated biological perspective. Science. 2014;345:45.CAS Article Google Scholar
- 18.Spikins P, Needham A, Wright B, Dytham C, Gatta M, Hitchens G. Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare. Quarternary Sci Rev. 2019;217:98–118.Article Google Scholar
- 19.Zwir I, Arnedo J, Del-Val C, Pulkki-Råback L, Konte B, Yang SS, et al. Three genetic-environmental networks for human personality. Mol Psychiatry 2019. https://doi.org/10.1038/s41380-019-0579-x. [Online ahead of print].
- 20.Lee PC, Fishlock V, Webber CE, Moss CJ. The reproductive advantages of a long life: longevity and senescence in wild female African Elephants. Behav Ecol Sociobiol. 2016;70:337–45.PubMed PubMed Central Article Google Scholar
- 21.Croft DP, Brent LJN, Franks DW, Cant MA. The evolution of long life after reproduction. Trends Ecol Evolut. 2015;30:407–16.Article Google Scholar
- 22.Hublin JJ. The origin of Neandertals. Proc Natl Acad Sci USA. 2009;106:16022–16027.CAS PubMed Article PubMed Central Google Scholar
- 23.Stringer C. The origin and evolution of Homo sapiens. Philos Trans R Soc Lond B Biol Sci. 2016;371:1–12.Article Google Scholar
- 24.Arsuaga JL, Martínez I, Arnold LJ, Aranburu A, Gracia-Téllez A, Sharp WD, et al. Neandertal roots: cranial and chronological evidence from Sima de los Huesos. Science. 2014;344:1358–63.CAS PubMed Article PubMed Central Google Scholar
- 25.Deino AL, Behrensmeyer AK, Brooks AS, Yellen JE, Sharp WD, Potts R. Chronology of the Acheulean to Middle Stone Age transition in eastern Africa. Science. 2018;360:95–98.CAS PubMed Article PubMed Central Google Scholar
- 26.Potts R, Behrensmeyer AK, Faith JT, Tryon CA, Brooks AS, Yellen JE, et al. Environmental dynamics during the onset of the Middle Stone Age in eastern Africa. Science. 2018;360:86–90.CAS PubMed Article PubMed Central Google Scholar
- 27.Owen RB, Muiruri VM, Lowenstein TK, Renaut RW, Rabideaux N, Luo S, et al. Progressive aridification in East Africa over the last half million years and implications for human evolution. Proc Natl Acad Sci USA. 2018;115:11174–11179.CAS PubMed Article PubMed Central Google Scholar
- 28.Marean CW, Bar-Matthews M, Bernatchez J, Fisher E, Goldberg P, Herries AIR, et al. Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature. 2007;449:905–908.CAS PubMed Article PubMed Central Google Scholar
- 29.Rito T, Vieira D, Silva M, Conde-Sousa E, Peira L, Mellars P, et al. A dispersal of Homo sapiens from southern to eastern Africa immediately preceded the out-of-Africa migration. Nat Sci Rep. 2019; 9.
- 30.Stiner MC, Kuhn SL. Changes in the “connectedness” and resilience of paleolithic societies in Mediterranean ecosystems. Hum Ecol. 2006;34:693–712.Article Google Scholar
- 31.Froehle AW, Churchill SE. Energetic competition between Neandertals and anatomically modern humans. Paleo Anthropology. 2009;2009:96–116.Google Scholar
- 32.Brumm A, Oktaviana AA, Burhan B, Hakim B, Lebe R, Zhao J-X, et al. Oldest cave art found in Sulawesi. Sci Adv. 2021;7:eabd4648.PubMed PubMed Central Article Google Scholar
- 33.Tierney JE, deMenocal PB, Zander PD. A climatic context for the out-of-Africa migration. Geology. 2017;45:1023–1026.Article Google Scholar
- 34.Stevenson A, Lindberg CA (eds). The New Oxford American Dictionary. Oxford University Press: New York, 2010.
- 35.Sternberg RJ. (ed). The nature of creativity. Cambridge University Press: Cambridge UK, 1988.
- 36.Sternberg RJ. A three-facet model of creativity. In: Sternberg RJ (ed). The nature of creativity. Cambridge University Press: Cambridge UK, 1988, pp 125–47.
- 37.Vernon PE (ed). Creativity. Penguin: Baltimore, 1970.
- 38.Sternberg RJ. The nature of creativity. Creativity Res J. 2006;18:87–98.Article Google Scholar
- 39.Guilford JP. Traits of creativity. In: Anderson HH (ed). Creativity and its cultivation. Harper: New York, 1959, pp 142-61.
- 40.Freud S. Creative writers and day-dreaming. In: Strachey J (ed). Standard edition of the complete psychological works of Sigmund Freud, vol. 9. Hogarth Press: London, 1908, pp 141–54.
- 41.Rogers CR. Towards a theory of creativity. ETC: A Rev Gen Semant. 1954;11:249–60.Google Scholar
- 42.Cloninger CR. Feeling good: the science of well-being. Oxford University Press: New York, 2004, 374pp.
- 43.Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci USA. 2001;98:676–82.CAS PubMed Article PubMed Central Google Scholar
- 44.Gusnard DA, Akbudak E, Shulman GL, Raichle ME. Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proc Natl Acad Sci USA. 2001;98:4259–64.CAS PubMed Article PubMed Central Google Scholar
- 45.Cloninger CR, Kedia S. The phylogenesis of human personality: Identifying the precursors of cooperation, altruism, & well-being. In: Sussman RW, Cloninger CR, (eds). The origins of cooperation and altruism. First edn. New York: Springer; 2011. p. 63–110.Google Scholar
- 46.Shen K-K, Welton T, Lyon M, McCorkidale AN, Sutherland GT, Burnham S, et al. Structural core of the executive control network: a high angle resolution diffusion MRI study. Hum Brain Mapp. 2020;41:1226–36.PubMed Article PubMed Central Google Scholar
- 47.Chavez-Eakle RA, Lara MDC, Cruz-Fuentes C. Personality: a possible bridge between creativity and psychopathology? Creativity Res J. 2006;18:27–38.Article Google Scholar
- 48.Takeuchi H, Taki Y, Sekiguchi A, Hashizume H, Nouchi R, Sassa Y, et al. Mean diffusivity of globus pallidus associated with verbal creativity measured by divergent thinking and creativity-related temperaments in young healthy adults. Hum Brain Mapp. 2015;36:1808–27.PubMed PubMed Central Article Google Scholar
- 49.Lieberman JN. Playfulness and divergent thinking: an investigation of their relationship at the kindergarten level. J Genet Psychol. 1965;107:219–24.CAS PubMed Article PubMed Central Google Scholar
- 50.Vosburg SK. The effects of positive and negative mood on divergent-thinking performance. Creativity Res J. 1998;11:165–72.Article Google Scholar
- 51.Cloninger CR, Svrakic DM, Przybeck TR. A psychobiological model of temperament and character. Arch Gen Psychiatry. 1993;50:975–90.CAS PubMed Article PubMed Central Google Scholar
- 52.Cloninger CR. What makes people healthy, happy, and fulfilled in the face of current world challenges? Mens Sana Monogr. 2013;11:16–24.PubMed PubMed Central Article Google Scholar
- 53.Cloninger CR, Cloniger KM. People create health: effective health promotion is a creative process. Int J Pers Cent Med. 2013;3:114–22.PubMed PubMed Central Google Scholar
- 54.Grucza RA, Goldberg LR. The comparative validity of 11 modern personality inventories: predictions of behavioral acts, informant reports, and clinical indicators. J Personal Assess. 2007;89:167–87.Article Google Scholar
- 55.Cloninger CR, Zohar AH. Personality and the perception of health and happiness. J Affect Disord. 2011;128:24–32.PubMed Article PubMed Central Google Scholar
- 56.Cloninger CR, Cloninger KM, Zwir I, Keltigangas-Jarvinen L. The complex genetics and biology of human temperament: a review of traditional concepts in relation to new molecular findings. Trans Psychiatry 2019.
- 57.Tulving E. Episodic memory and common sense: how far apart? Philos Trans R Soc Lond B Biol Sci. 2001;356:1505–15.CAS PubMed PubMed Central Article Google Scholar
- 58.Ambrose SH. Coevolution of composite-tool technology, constructive memory, and language: implicatiions for the evolution of modern human behavior. Curr Anthropol. 2010;51:135–47.Article Google Scholar
- 59.Spreng RN, Mar RA, Kim ASN. The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: a quantitative meta-analysis. J Cogn Neurosci. 2008;21:489–510.Article Google Scholar
- 60.Cicchetti D, Rogosch FA. Equifinality and multifinality in developmental psychopatology. Dev Psychopathol. 1996;8:597–600.Article Google Scholar
- 61.Wright S. The shifting balance theory and macroevolution. Annu Rev Genet. 1982;16:1–19.CAS PubMed Article PubMed Central Google Scholar
- 62.Wang Z, Liao B-Y, Jianzhi Z. Genomic patterns of pleiotropy and the evolution of complexity. Proc Natl Acad Sci USA. 2010;107:18034–18039.CAS PubMed Article PubMed Central Google Scholar
- 63.Hill WG, Zhang XS. On the pleiotropic structure of the genotype-phenotype map and the evolvability of complex organisms. Genetics. 2012;190:1131–1137.PubMed PubMed Central Article Google Scholar
- 64.Zwir I, Arnedo J, Del-Val C, Pulkki-Raback L, Konte B, Yang SS, et al. Uncovering the complex genetics of human character. Mol Psychiatry. 2020;25:2295–312.CAS PubMed Article PubMed Central Google Scholar
- 65.Zwir I, Arnedo J, Del-Val C, Pulkki-Raback L, Konte B, Yang SS, et al. Uncovering the complex genetics of human temperament. Mol Psychiatry. 2020;25:2275–94.CAS PubMed Article PubMed Central Google Scholar
- 66.Gould SJ, Vrba ES. Exaptation—a missing term in the science of form. Paleobiology. 1982;8:4–15.Article Google Scholar
- 67.Hauser MD, Chomsky N, Fitch WT. The faculty of language: what is it, who has it, and how did it evolve? Science. 2002;298:1569–79.CAS PubMed Article PubMed Central Google Scholar
- 68.McBrearty S, Brooks AS. The revolution that wasn’t: a new interpretation of the origin of modern human behavior. J Hum Evol. 2000;39:453–563.CAS PubMed Article PubMed Central Google Scholar
- 69.Pearce E, Stringer C, Dunbar RI. New insights into differences in brain organization between Neanderthals and anatomically modern humans. Proc Biol Sci. 2013;280:20130168.PubMed PubMed Central Google Scholar
- 70.Bruner E, Ogihara N, Tanabe HC (eds). Digital endocasts: from skulls to brains. Springer: Tokyo, 2018.
- 71.Akazawa T, Ogihara N, Tanabe HC, Terashima H (eds). Dynamics of learning in Neanderthals and modern humans: cognitive and physical perspectives. Springer: Tokyo, 2014.
- 72.Soressi M, McPherron SP, Lenoir M, Dogandzic T, Goldberg P, Jacobs Z, et al. Neandertals made the first specialized bone tools in Europe. Proc Natl Acad Sci USA. 2013;110:14186–90.CAS PubMed Article PubMed Central Google Scholar
- 73.Radovcic D, Srsen AO, Radovcic J, Frayer DW. Evidence for Neandertal jewelry: modified white-tailed eagle claws at Krapina. PLoS One. 2015;10:e0119802.PubMed PubMed Central Article CAS Google Scholar
- 74.Hoffmann DL, Standish CD, Garcia-Diez M, Pettitt PB, Milton JA, Zilhao J, et al. U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art. Science. 2018;359:912–915.CAS PubMed Article PubMed Central Google Scholar
- 75.Rodriguez-Vidal J, d’Errico F, Giles Pacheco F, Blasco R, Rosell J, Jennings RP, et al. A rock engraving made by Neanderthals in Gibraltar. Proc Natl Acad Sci USA. 2014;111:13301–13306.CAS PubMed Article PubMed Central Google Scholar
- 76.Kolbert E. The madness gene: homo neanderthalensis. The sixth extinction: an unnatural history. Picador: New York, 2014, p 248.
- 77.DeSalle R, Tattersall I. What aDNA can (and cannot) tell us about the emergence of language and speech. J Lang Evolut. 2018;3:59–66.Article Google Scholar
- 78.Quoidbach J, Hansenne M, Mottet C. Personality and mental time travel: a differential approach to autonoetic consciousness. Conscious Cognition. 2008;17:1082–92.Article Google Scholar
- 79.Van Schuerbeek P, Baeken C, De Raedt R, De Mey J, Luypaert R. Individual differences in local gray and white matter volumes reflect differences in temperament and character: a voxel-based morphometry study in healthy young females. Brain Res. 2011;1371:32–42.PubMed Article CAS PubMed Central Google Scholar
- 80.Zohar AH, Cloninger CR, McCraty R. Personality and heart rate variability: exploring pathways from personality to cardiac coherence and health. J Soc Sci. 2013;1:32–39.Google Scholar
- 81.Zwir I, Mishra P, Del-Val C, Gu CC, de Erausquin G, Lehtimäki T, et al. Uncovering the complex genetics of human personality: response from authors on the PGMRA model. Mol Psychiatry. 2019.
- 82.Moreira PAS, Inman RA, Cloninger CR. Personality networks and behavioral problems: Integrating temperament and character using latent profile and latent class analysis. Child Psychiatry Hum Dev. 2020;2020:1–13.Google Scholar
- 83.Raitakari OT, Juonala M, Ronnemaa T, Keltikangas-Jarvinen L, Rasanen L, Pietikainen M, et al. Cohort profile: the cardiovascular risk in Young Finns Study. Int J Epidemiol. 2008;37:1220–1226.PubMed Article PubMed Central Google Scholar
- 84.Ravaja N, Keltikangas-Jarvinen K. Cloninger’s temperament and character dimensions in young adulthood and their relation to characteristics of parental alcohol use and smoking. J Stud Alcohol. 2001;62:98–104.CAS PubMed Article PubMed Central Google Scholar
- 85.Yates A, Akanni W, Amode MR, Barrell D, Billis K, Carvalho-Silva D, et al. Ensembl 2016. Nucleic Acids Res. 2016;44:D710–716.CAS PubMed Article PubMed Central Google Scholar
- 86.Smedley D, Haider S, Durinck S, Pandini L, Provero P, Allen J, et al. The BioMart community portal: an innovative alternative to large, centralized data repositories. Nucleic Acids Res. 2015;43:W589–598.CAS PubMed PubMed Central Article Google Scholar
- 87.Aken BL, Achuthan P, Akanni W, Amode MR, Bernsdorff F, Bhai J, et al. Ensembl 2017. Nucleic Acids Res. 2017;45:D635–D642.CAS PubMed Article PubMed Central Google Scholar
- 88.Huang W, Tsai L, Li Y, Hua N, Sun C, Wei C. Widespread of horizontal gene transfer in the human genome. BMC Genom. 2017;18:274.Article CAS Google Scholar
- 89.K Prüfer C Filippo SG Mafessoni F Korlević PM Hajdinjak et al. 2017 A high-coverage Neandertal genome from Vindija Cave in Croatia Science 358 655 658.PubMed PubMed Central Article CAS Google Scholar
- 90.Mafessoni F, Grote S, de Filippo C, Slon V, Kolobova KA, Viola B, et al. A high-coverage Neandertal genome from Chagyrskaya Cave. Proc Natl Acad Sci USA. 2020;117:15132–15136.CAS PubMed Article PubMed Central Google Scholar
- 91.Cohen J. Statistical power analysis for the behavioral sciences. 2nd edn. Lawrence Erlbaum: Hillsdale, NJ, 1988.
- 92.Zhang Z, Yuan K-H. Practical statistical power analysis using Webpower and R. ISDSA Press: Granger, IN, 2018, pp. 356 .
- 93.Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, et al. Initial sequencing and analysis of the human genome. Nature. 2001;409:860–921.CAS PubMed Article PubMed Central Google Scholar
- 94.1000 Genomes Project C, Abecasis GR, Altshuler D, Auton A, Brooks LD, Durbin RM, et al. A map of human genome variation from population-scale sequencing. Nature. 2010;467:1061–73.Article CAS Google Scholar
- 95.Ward LD, Kellis M. Evidence of abundant purifying selection in humans for recently acquired regulatory functions. Science. 2012;337:1675–1678.CAS PubMed PubMed Central Article Google Scholar
- 96.Biswas S, Akey JM. Genomic insights into positive selection. Trends Genet. 2006;22:437–46.CAS PubMed Article PubMed Central Google Scholar
- 97.Poblete GF, Gosnell SN, Meyer M, Fang M, Nyugen T, Patriquin MA, et al. Process genes list: an approach to link genetics and human brain imaging. J Neurosci Methods. 2020; 339.
- 98.Mineka S, Suomi SJ. Social separation in monkeys. Psychol Bull. 1978;85:1376–1400.CAS PubMed Article PubMed Central Google Scholar
- 99.Bowlby J. Attachment. Second edn. Basic Books: New York, 1983.
- 100.de Waal FB. Primates–a natural heritage of conflict resolution. Science. 2000;289:586–90.PubMed Article PubMed Central Google Scholar
- 101.de Waal FB. Cultural primatology comes of age. Nature. 1999;399:635–636.PubMed Article CAS PubMed Central Google Scholar
- 102.Whiten A, Spiteri A, Horner V, Bonnie KE, Lambeth SP, Schapiro SJ, et al. Transmission of multiple traditions within and between chimpanzee groups. Curr Biol. 2007;17:1038–43.CAS PubMed Article PubMed Central Google Scholar
- 103.Sussman RW, Chapman AR (eds). The origins and nature of sociality. Aldine de Gruyter: New York, 2004.
- 104.de Waal FB. Macaque social culture: development and perpetuation of affiliative networks. J Comp Psychol. 1996;110:147–54.PubMed Article PubMed Central Google Scholar
- 105.Suddendorf T, Whiten A. Mental evolution and development: evidence for secondary representation in children, great apes, and other animals. Psychol Bull. 2001;127:629–50.CAS PubMed Article PubMed Central Google Scholar
- 106.Donald M. Origins of the modern mind: three stages in the evolution of culture and cognition. Harvard University Press: Cambridge, MA, 1991.
- 107.Povinelli DJ. Folk physics for apes: the chimpanzee’s theory of how the world works. Oxford University Press: New York, 2000.
- 108.Povinelli DJ, Giambrone S. Reasoning about beliefs: a human specialization? Child Dev. 2001;72:691–695.CAS PubMed Article PubMed Central Google Scholar
- 109.McPherron SP, Alemseged Z, Marean CW, Wynn JG, Reed D, Geraads D, et al. Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature. 2010;466:857–60.CAS PubMed Article PubMed Central Google Scholar
- 110.Castellano S, Parra G, Sanchez-Quinto FA, Racimo F, Kuhlwilm M, Kircher M, et al. Patterns of coding variation in the complete exomes of three Neandertals. Proc Natl Acad Sci USA. 2014;111:6666–71.CAS PubMed Article PubMed Central Google Scholar
- 111.Niven L, Steele TE, Rendu W, Mallye JB, McPherron SP, Soressi M, et al. Neandertal mobility and large-game hunting: the exploitation of reindeer during the Quina Mousterian at Chez-Pinaud Jonzac (Charente-Maritime, France). J Hum Evol. 2012;63:624–35.PubMed Article PubMed Central Google Scholar
- 112.Sandgathe DM, Dibble HL, Goldberg P, McPherron SP, Turq A, Niven L, et al. Timing of the appearance of habitual fire use. Proc Natl Acad Sci USA. 2011;108:E298. author reply E299CAS PubMed Article PubMed Central Google Scholar
- 113.Tattersall I. The world from beginnings to 4000 BCE. Oxford University Press: New York, 2008b.
- 114.Sandgathe DM, Dibble HL, Goldberg P, McPherron SP. The Roc de Marsal Neandertal child: a reassessment of its status as a deliberate burial. J Hum Evol. 2011;61:243–53.PubMed Article PubMed Central Google Scholar
- 115.Hagen MA. Development of ability to perceive and produce pictorial depth cue of overlapping. Percept Mot Skills. 1976;42:1007–14.CAS PubMed Article PubMed Central Google Scholar
- 116.Prufer K, Racimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, et al. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature. 2014;505:43–49.PubMed Article CAS PubMed Central Google Scholar
- 117.Green RE, Krause J, Briggs AW, Maricic T, Stenzel U, Kircher M, et al. A draft sequence of the Neandertal genome. Science. 2010;328:710–22.CAS PubMed PubMed Central Article Google Scholar
- 118.Brooks KR. Depth perception and the history of three-dimensional art: who produced the first stereoscopic images? Iperception. 2017;8:2041669516680114.PubMed PubMed Central Google Scholar
- 119.Fu Q, Li H, Moorjani P, Jay F, Slepchenko SM, Bondarev AA, et al. Genome sequence of a 45,000-year-old modern human from western Siberia. Nature. 2014;514:445–449.CAS PubMed PubMed Central Article Google Scholar
- 120.Levine B. Autobiographical memory and the self in time: brain lesion effects, functional neuroanatomy, and lifespan development. Brain Cognition. 2004;55:54–68.PubMed Article PubMed Central Google Scholar
- 121.Moll J, Krueger F, Zahn R, Pardini M, de Oliveira-Souza R, Grafman J. Human fronto-mesolimbic networks guide decisions about charitable donation. Proc Natl Acad Sci USA. 2006;103:15623–15628.CAS PubMed Article PubMed Central Google Scholar
- 122.Ulitsky I. Evolution to the rescue: using comparative genomics to understand long non-coding RNAs. Nat Rev Genet. 2016;17:601–614.CAS PubMed Article PubMed Central Google Scholar
- 123.Fang Y, Fullwood MJ. Roles, functions, and mechanisms of long non-coding RNAs in cancer. Genomics Proteom Bioinforma. 2016;14:42–54.Article Google Scholar
- 124.Quinn JJ, Chang HY. Unique features of long non-coding RNA biogenesis and function. Nat Rev Genet. 2016;17:47–62.CAS PubMed Article PubMed Central Google Scholar
- 125.Lea A, Subramaniam M, Ko A, Lehtimäki T, Raitoharju E, Kahonen M, et al. Genetic and environmental perturbations lead to regulatory decoherence. eLife. 2019;8:e40538.PubMed PubMed Central Article Google Scholar
- 126.Rilling JK, Goldsmith DR, Glenn AL, Jairam MR, Elfenbein HA, Dagenais JE, et al. The neural correlates of the affective response to unreciprocated cooperation. Neuropsychologia. 2008;46:1256–1266.PubMed Article PubMed Central Google Scholar
- 127.Coxworth JE, Kim PS, McQueen JS, Hawkes K. Grandmothering life histories and human pair bonding. Proc Natl Acad Sci USA. 2015;112:11806–11811.CAS PubMed Article PubMed Central Google Scholar
- 128.Hawkes K, O’Connell JF, Jones NG, Alvarez H, Charnov EL. Grandmothering, menopause, and the evolution of human life histories. Proc Natl Acad Sci USA. 1998;95:1336–1339.CAS PubMed Article PubMed Central Google Scholar
- 129.Kim PS, Coxworth JE, Hawkes K. Increased longevity evolves from grandmothering. Proc Biol Sci. 2012;279:4880–4884.PubMed PubMed Central Google Scholar
- 130.Hawkes K, Kim PS, Kennedy B, Bohlender R, Hawks J. A reappraisal of grandmothering and natural selection. Proc Biol Sci. 2011;278:1936–1938. discussion 1939-1941PubMed PubMed Central Google Scholar
- 131.Schwarz F, Springer SA, Altheide TK, Varki NM, Gagneux P, Varki A. Human-specific derived alleles of CD33 and other genes protect against postreproductive cognitive decline. Proc Natl Acad Sci USA. 2016;113:74–79.CAS PubMed Article PubMed Central Google Scholar
- 132.Tattersall I. Brain size and the emergence of modern human cognition In: Schwartz JH (ed). Rethinking human evolution, First edn. MIT Press: Cambridge, MA, 2018, pp 319-334.
- 133.Scally A. The mutation rate in human evolution and demographic inference. Curr Opin Genet Dev. 2016;41:36–43.CAS PubMed Article PubMed Central Google Scholar
- 134.Kaessmann H. Origins, evolution, and phenotypic impact of new genes. Genome Res. 2010;20:1313–1326.CAS PubMed PubMed Central Article Google Scholar
- 135.King MC, Wilson AC. Evolution at two levels in humans and chimpanzees. Science. 1975;188:107–116.CAS PubMed Article PubMed Central Google Scholar
- 136.Carroll SB. Evolution at two levels: on genes and form. PLoS Biol. 2005;3:e245.PubMed PubMed Central Article CAS Google Scholar
- 137.Gilad Y, Oshlack A, Rifkin SA. Natural selection on gene expression. Trends Genet. 2006;22:456–461.CAS PubMed Article PubMed Central Google Scholar
- 138.Ji Z, Song R, Regev A, Struhl K. Many lncRNAs, 5’UTRs, and pseudogenes are translated and some are likely to express functional proteins. Elife. 2015;4:e08890.PubMed PubMed Central Article Google Scholar
- 139.Crisp A, Boschetti C, Perry M, Tunnacliffe A, Micklem G. Expression of multiple horizontally acquired genes is a hallmark of both vertebrate and invertebrate genomes. Genome Biol. 2015;16:50.PubMed PubMed Central Article Google Scholar
- 140.Schrider DR, Navarro FC, Galante PA, Parmigiani RB, Camargo AA, Hahn MW, et al. Gene copy-number polymorphism caused by retrotransposition in humans. PLoS Genet. 2013;9:e1003242.CAS PubMed PubMed Central Article Google Scholar
- 141.Navarro FC, Galante PA. A genome-wide landscape of retrocopies in primate genomes. Genome Biol Evol. 2015;7:2265–2275.CAS PubMed PubMed Central Article Google Scholar
- 142.Barbash S, Simchovitz A, Buchman AS, Bennett DA, Shifman S, Soreq H. Neuronal-expressed microRNA-targeted pseudogenes compete with coding genes in the human brain. Transl Psychiatry. 2017;7:e1199.CAS PubMed PubMed Central Article Google Scholar
- 143.Kaneko T, Tomonaga M. The perception of self-agency in chimpanzees (Pan trogloytes). Proc R Soc B. 2011;278:3694–3702.PubMed Article PubMed Central Google Scholar
- 144.Povinelli DJ. Can comparative psychology crack its toughest nut? Anim Behav Cognition. 2020;7:589–652.Article Google Scholar
- 145.Zohar AH, Zwir I, Wang J, Cloninger CR, Anokhin AP. The development of temperament and character during adolescence: the processes and phases of change. Dev Psychopathol. 2019;31:601–617.PubMed Article PubMed Central Google Scholar
Affiliations
- Department of Psychiatry, Washington University School of Medicine, St. Louis, MO, USAI. Zwir & C. R. Cloninger
- Department of Computer Science and Artificial Intelligence, University of Granada, Andalusian Research Institute in Data Science and Computational Intelligence, Granada, SpainI. Zwir, C. Del-Val, R. Romero-Zaliz, A. Mesa & J. Arnedo
- Unit of Psychology, Faculty of Education, University of Oulu, Oulu, FinlandM. Hintsanen
- Anthropedia Foundation, St. Louis, MO, USAK. M. Cloninger & C. R. Cloninger
- The Menninger Clinic, Baylor College of Medicine, and DeBakey VA Medical Center, Houston, TX, USAR. Salas & G. F. Poblete
- The Menninger Clinic, Houston, TX, USAG. F. Poblete
- Department of Clinical Chemistry, Fimlab Laboratories, and Finnish Cardiovascular Research Center – Tampere, Faculty of Medicine and Health Technology, Tampere University, Tampere, FinlandE. Raitoharju & T. Lehtimäki
- Center for Population Health Research, University of Turku and Turku University Hospital; Research Center of Applied and Preventive Cardiovascular Medicine, University of Turku; Department of Clinical Physiology and Nuclear Medicine, Turku University Hospital, Turku, FinlandO. Raitakari
- Department of Psychology and Logopedics, University of Helsinki, Helsinki, FinlandL. Keltikangas-Järvinen
- Department of Psychiatry, University of Texas San Antonio, Long School of Medicine, The Glenn Briggs Institute of Alzheimer’s and Neurodegenerative Disorders, San Antonio, TX, USAG. A. de Erausquin
- American Museum of Natural History, New York, NY, USAI. Tattersall





Mille mercis pour cette traduction car je m’apprêtais à traduire l’étude par Google automatique. Gratitude donc !
Un commentaire qui fait plaisir !